Back Clinic қартаюға қарсы хиропрактикалық және функционалдық медицина тобы. Біздің денеміз өмір сүру үшін үздіксіз және ешқашан бітпейтін шайқаста. Жасушалар туады, жасушалар жойылады. Ғалымдар әрбір жасуша реактивті оттегі түрлерінің (ROS) немесе бос радикалдардың 10,000 XNUMX-нан астам жеке шабуылдарына төтеп беруі керек деп есептейді. «Үздіксіз» денеде шабуылға төтеп беретін және зақымдалған немесе жойылған нәрсені қалпына келтіретін өзін-өзі емдеудің керемет жүйесі бар. Бұл дизайнымыздың сұлулығы.
Қартаю биологиясын түсіну және ғылыми түсініктерді емдеу арқылы кейінгі өмірдің денсаулығын жақсартатын араласуларға аудару. Қартаюға қарсы емнің нақты нені құрайтыны туралы нақты, консенсус көзқарасы болуы пайдалы.
Понсе де Леонның ұзақ өмір сүруге ұмтылған күндерінен бұрын адам мәңгілік жастық шаққа арбады. Хиропрактикалық күтім оның денсаулық қозғалысымен өзін-өзі емдеу қабілетін тұрақтандырудың және жақсартудың күшті әдісі болып табылады. Доктор Алекс Хименес қартаюға қарсы пандораға қатысты тұжырымдамаларды талқылайды.
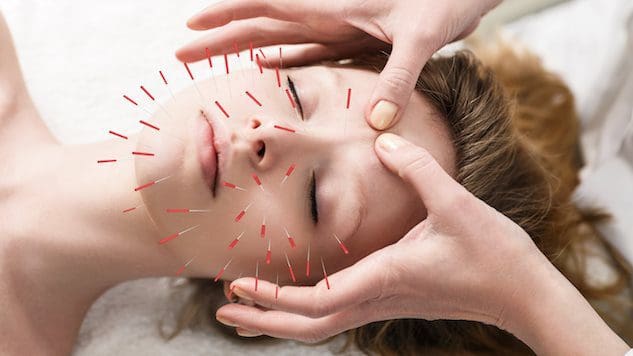
Терінің денсаулығын жақсартқысы келетін немесе сақтағысы келетін адамдар үшін акупунктураны қосу теріні жақсартуға және қартаю процесімен күресуге көмектесе ала ма?
Косметикалық акупунктура
Косметикалық акупунктура инені енгізудің дәстүрлі акупунктура тәжірибесіне сәйкес келеді. Мақсат - қартаю белгілерін жою және тері денсаулығын жақсарту. Оны кейде бетті инемен жасару деп атайды, ол хирургиялық бет көтеруге және басқа да дәстүрлі процедураларға балама ретінде қолданылған. Алдын ала зерттеулер оның қартаю дақтарын кетіруге, салбыраған қабақтарды көтеруге және әжімдерді азайтуға қалай көмектесетінін зерттеді. (Younghee Yun және т.б., 2013 ж)
Акупунктура қалай жұмыс істейді
Дәстүрлі қытай медицинасында немесе TCM-де акупунктура бүкіл денеде энергия ағынын - qi немесе chi - жақсарту үшін бұрыннан қолданылған. Бұл энергия меридиандар деп аталатын энергетикалық жолдар арқылы айналады деп саналады. Денсаулыққа қатысты проблемалар туындаған кезде, TCM мәліметтері бойынша, айналымда кедергілер немесе бітелулер бар.
Акупунктур мамандары оңтайлы қан айналымын/ағынды қалпына келтіріп, инелерді арнайы нүктелерге енгізу арқылы денсаулықты жақсарта алады. (Ұлттық денсаулық институттары, 2007 ж)
Косметикалық акупунктура
Косметикалық акупунктура терінің денсаулығын жақсартады және коллаген өндірісін ынталандыру арқылы қартаюға қарсы ем ретінде әрекет етеді. Бұл ақуыз терінің негізгі құрамдас бөлігі болып табылады. Терінің ішкі қабаты дене қартаюына қарай коллаген мен қаттылығын жоғалтады. Дегенмен, акупунктура коллаген өндірісіне ықпал ете алады деген мәлімдемені растау үшін қосымша зерттеулер қажет. Кейбіреулер косметикалық акупунктура дененің жалпы энергиясын жақсарту арқылы теріні жасартуға көмектеседі деп болжайды. Бір зерттеуде адамдар бет косметикалық акупунктурасының бес сеансынан кейін жақсартуларды байқаған. (Younghee Yun және т.б., 2013 ж) Дегенмен оңтайлы нәтижеге жету үшін аптасына бір немесе екі рет он процедураны орындау ұсынылады. Осыдан кейін күтім процедуралары әр төрт-сегіз апта сайын жасалады. Ботокс немесе тері толтырғыштарынан айырмашылығы, косметикалық акупунктура тез шешілмейді. Негізгі назар тері мен денеде ұзақ мерзімді өзгерістерді жасау болып табылады, бұл жақсаруды білдіреді:
Инелер теріге енгізілгенде, олар оң микротравмалар деп аталатын жараларды жасайды. Дененің табиғи емдеу және қалпына келтіру қабілеттері бұл жараларды сезген кезде белсендіріледі. Бұл пункциялар тері жасушаларына қоректік заттар мен оттегіні жеткізетін лимфа және қан айналымы жүйелерін ынталандырады, оларды ішінен сыртқа тамақтандырады.
Бұл бет терісін тегістеуге көмектеседі және терінің жарқырауына ықпал етеді.
Оң микротравмалар да коллаген өндірісін ынталандырады.
Бұл серпімділікті жақсартуға, сызықтар мен әжімдерді азайтуға көмектеседі.
баламалары
Бірнеше табиғи құралдар тері денсаулығын жақсартуға және қартаюға қарсы артықшылықтарды ұсынуға көмектеседі. Керамидтер – терінің үстіңгі қабатында табиғи түрде кездесетін май молекуласы және тері күтімі өнімдерінде қолданылатын ингредиент. Олар терідегі қартаюға байланысты құрғақтықтан қорғай алады. (Л Ди Марцио 2008 ж) Алдын ала зерттеулер ақ шайды теріге жағу коллаген мен эластиннің – терінің серпімділігін қолдайтын және салбырауды болдырмайтын ақуыздың ыдырауымен күресуі мүмкін екенін көрсетеді). Сондай-ақ, арган майы, бұршақ майы және теңіз шырғанағы сияқты табиғи заттардың теріні жақсартатын ылғалдандыратын артықшылықтарды ұсына алатындығы туралы дәлелдер бар.(Tamsyn SA Thring et al., 2009)
Косметикалық акупунктураның қосымша дәлелдері қажет болғанымен, акупунктураны біріктіру стрессті басқаруға және жалпы денсаулықты жақсартуға көмектеседі. Косметикалық акупунктураны қарастыратын адамдар бұл оларға сәйкес келетінін білу үшін бастапқы медициналық көмек көрсетушімен кеңесу керек.
Денсаулықты бірге нығайту: көпсалалы бағалау мен емдеуді қабылдау
Әдебиеттер тізімі
Юн, Ю., Ким, С., Ким, М., Ким, К., Пак, Дж.С., Чой, И. (2013). Бет косметикалық акупунктурасының бет икемділігіне әсері: ашық жапсырма, бір қолды пилоттық зерттеу. Дәлелдерге негізделген қосымша және балама медицина: eCAM, 2013, 424313. doi.org/10.1155/2013/424313
Куге, Х., Мори, Х., Танака, Т.Х. және Цудзи, Р. (2021). Бет-әлпетті тексеру парағының (FCS) сенімділігі мен жарамдылығы: косметикалық акупунктурамен өзін-өзі қанағаттандыру үшін бақылау тізімі. Дәрілік заттар (Базель, Швейцария), 8(4), 18. doi.org/10.3390/medicines8040018
Di Marzio, L., Cinque, B., Cupelli, F., De Simone, C., Cifone, MG, & Giuliani, M. (2008). Streptococcus thermophilus бактериялық сфингомиелиназасын қысқа мерзімді жергілікті қолданудан кейін қарт адамдарда тері-керамид деңгейінің жоғарылауы. Иммунопатология және фармакология халықаралық журналы, 21(1), 137–143. doi.org/10.1177/039463200802100115
Thring, TS, Hili, P., & Naughton, DP (2009). 21 өсімдік сығындыларының коллагеназаға қарсы, эластазаға қарсы және антиоксиданттық белсенділігі. BMC қосымша және балама медицина, 9, 27. doi.org/10.1186/1472-6882-9-27
Адамның омыртқасын жоғарғы пішінде ұстау ауырсынуды азайтуға және көбірек қозғалғыштыққа, икемділікке және еркіндікке тең. Дене тозып кетеді және бұл әрқайсымызда болатын қартаюдың табиғи әсері. Қартаюға байланысты омыртқа мәселелері жаттығулармен, созылумен және хиропротикалық күтіммен шешілмесе және шешілмесе, күрделі болуы мүмкін.
Қартаю және арқа
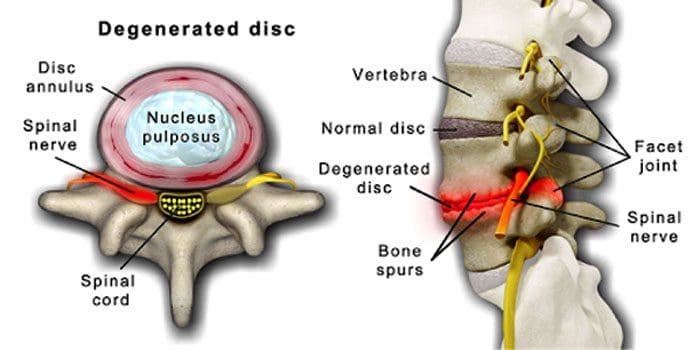
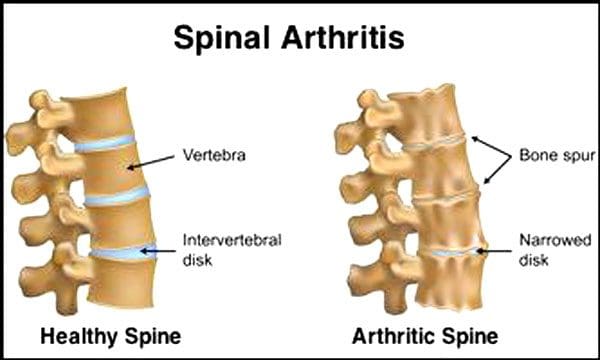
Жұлынның дискілері мен буындарының жасына қарай нашарлауы қалыпты жағдай. Омыртқаның стенозы немесе жұлын арнасының тарылуы да қартаю процесінің бір бөлігі болуы мүмкін. Қартаюдың екі шарты бар дискреативті диск ауруы және артрит бұл да қамтуы мүмкін омыртқаның байламдарының қатайуы және остеопороз.
Дегенеративті диск ауруы 40 жастағы адамдардың 40% -ында кездеседі
80 жастан асқан адамдар үшін 80%-ға дейін артады.
Ол айналада орналасқан бірте-бірте негізінен судан негізінен майға ауысатын дискілер.
Майлы болған кезде дискілер тарылып, серпімділігін жоғалтады.
Бұл туралы Ауруларды бақылау және алдын алу орталықтары хабарлады Американдық ересектердің 23% артритпен ауырады. Бұл негізінен фасеттік буындарға әсер ететін жағдай. Буындар ісінеді, бұл қозғалыс ауқымын азайтады және жұлын нервтеріне әсер етуі мүмкін, ауырсынуды, әлсіздікті және сіатиканы тудырады. Уақыт өте омыртқаның айналасындағы және ішіндегі байламдар қатайып, қозғалыс ауқымын азайтып, стенозды тудырады.. Сүйек жоғалуы немесе остеопороз гормондардың өзгеруіне және тамақтану сияқты басқа факторларға байланысты. Қартаю - бұл табиғи процесс, бірақ адамдар қанша жаста болса да, омыртқаларының жоғары пішінде болуына көмектесе алады.
Салауатты қалыпқа жаттықтыру
Ботадан бірден сау дененің дұрыс механикасы міндетті болып табылады. Дене қалпын білу және есте ұстау денені теңестіруді және теңгерімді сақтайды. Салауатты поза келесі әсерлерді азайтуға көмектеседі:
Жұлынның стенозы
Диегеративті диск ауруы
Гернация
Омыртқаның сыну қаупі
Дұрыс қалып жаттықтыру мыналарды қамтиды:
Еңкейуді азайтыңыз
Жұмыс станциясының жоғарғы пішінде және эргономикалық тұрғыдан дұрыс екеніне көз жеткізіңіз
Көтеру кезінде тізе бүгіп, омыртқаны мүмкіндігінше тік ұстаңыз.
Йога
Йога сау, жас омыртқа үшін өте пайдалы болуы мүмкін. Йога омыртқаны жоғары пішінде ұстау үшін үш бағытты орындайды. Бұл мыналарды қамтиды:
Тұрақты жаттығу
Икемділікті сақтайды
Идеал дене салмағына жетеді
Йога - бұл омыртқа үшін жасқа қарсы әрекет. Өйткені ол:
Күшті сақтайды
икемділік
Қалып
баланс
Әртүрлі омыртқа жағдайларына, әсіресе артрит ауруы үшін пайдалы болуы мүмкін
Құлаулар ауыр жарақаттарға әкелуі мүмкін. Йога те тепе-теңдікте жұмыс істеуге көмектеседі.
Хиропрактикке барыңыз
Профилактикалық медицина денені сау, жас және мүмкіндігінше күшті ұстаудың кілті болып табылады. Хиропрактикалық тексеру омыртқаның проблемалары бар-жоғын және оңтайлы емдеу жоспарын әзірлеу үшін диагнозды анықтай алады. Арқадағы және/немесе аяқтың ауырсынуына байланысты дене қызметі шектелген болса, Injury Medical Chiropractic and Functional Medicine клиникасына хабарласыңыз және омыртқаны жоғары пішінге келтіріңіз.
Дене құрамы
Жаттығу/Тұрақтылық шарының бұйралары
Бұл жаттығу Жұлын күшіне тән бұлшықет топтарын жұмыс істейді және мыналарды қамтиды:
бицепсы бедер
Глиталар
Терең іш
Жамбас ұрлағыштары мен ротаторлары
Мұндай жаттығулар - бұл буындардағы, жамбастардағы функционалдық күш пен төзімділікті арттырудың және жарақаттардың алдын алудың ең тиімді әдістерінің бірі. Бұл жаттығуды орындау үшін:
Тізеңізді бүгіп, арқаңызда жатыңыз
Аяқтарды жоғары көтеріңіз, аяқтың төменгі бөлігі жаттығу допының үстіне тіреледі
Аяқтарыңызды түзу болғанша айналдырыңыз
Позицияны бір-екі секунд ұстаңыз
Аяқтарды қысу кезінде қозғалыстың жоғарғы жағына оралыңыз
Бұл бұлшықеттермен жұмыс істеу омыртқадағы иілу, иілу немесе иілу қозғалыстарын жеңілдетеді.
Доктор Алекс Хименестің блогындағы хабарламадан бас тарту
Біздің ақпарат шеңберіміз хиропрактикалық, тірек-қимыл аппараты, физикалық дәрі-дәрмектермен, сауықтыру және денсаулыққа қатысты мәселелермен және / немесе функционалды медицина туралы мақалалармен, тақырыптармен және пікірталастармен шектеледі. Біз тірек-қимыл аппаратының жарақаттары немесе бұзылулары кезінде емдеуді қолдау және қолдау үшін функционалды денсаулық және сауықтыру хаттамаларын қолданамыз. Біздің жазбаларымыз, тақырыптарымыз, тақырыптарымыз бен түсініктеріміз клиникалық мәселелерге тікелей немесе жанама байланысты және қолдау көрсететін клиникалық мәселелерді, мәселелер мен тақырыптарды қамтиды. *
Біздің кеңсе қолдау көрсететін дәйексөздер беруге негізделген талпыныстар жасады және біздің зерттеулерімізді немесе зерттеулерімізді анықтады. Біз сондай-ақ көмекші зерттеулердің көшірмелерін кеңестің және қоғамның сұрауы бойынша қол жетімді етеміз. Біз оның белгілі бір күтім жоспарына немесе емдеу хаттамасына қалай көмектесуі мүмкін екендігі туралы қосымша түсініктеме талап ететін мәселелерді қарастыратынымызды түсінеміз; сондықтан жоғарыда аталған тақырыпты одан әрі талқылау үшін доктор Алекс Хименестен немесе 915-850-0900 нөмірі бойынша бізбен байланысыңыз. Техас және Нью-Мексикода лицензияланған провайдерлер (лер) *
Біз тұтынатын тағамдардың денсаулығымызға пайдалы немесе зиянды болуы мүмкін. Нашар тамақтану денсаулықтың әртүрлі мәселелерін, соның ішінде семіздік, жүрек-қан тамырлары аурулары және 2 типті қант диабетін тудыруы мүмкін. Сонымен қатар, дұрыс тамақтану сізді қуаттандырады, денсаулыққа қатысты мәселелер қаупін азайтады, сонымен қатар салауатты салмақты сақтауға және реттеуге көмектеседі. Ұзақ өмір сүруді қаласаңыз, денеңізді жақсы тағамдармен толтыруыңыз керек. Келесі мақалада біз жалпы денсаулық пен денсаулықты жақсартуға көмектесетін ұзақ өмір сүруге көмектесетін бірнеше жақсы тағамдарды тізімдейміз.
Крест тәрізді көкөністер
Крестгүлді көкөністер біздің гормондарымызды өзгертуге, дененің табиғи детоксикация жүйесін іске қосуға және тіпті ісік жасушаларының өсуін азайтуға бірегей қабілеті бар. Пайдалы қасиеттерін босату үшін оларды жақсылап шайнау немесе ұсақтап, туралған, шырындары немесе араластырып жеу керек. Айқышгүлді көкөністерде кездесетін сульфорафан қан тамырларының қабырғаларын жүрек ауруын тудыруы мүмкін қабынудан қорғауға көмектесетіні анықталды. Қырыққабат, қырыққабат, Брюссель өскіндері, гүлді қырыққабат және брокколи сияқты крестгүлді көкөністер әлемдегі қоректік заттарға ең бай тағамдардың бірі болып табылады.
Жасыл салаттар
Шикі жапырақты жасылдарда бір фунт үшін 100 калориядан аз, бұл оларды салмақ жоғалту үшін тамаша тағам етеді. Көбірек жасыл салаттарды жеу сонымен қатар инфаркт, инсульт, қант диабеті және қатерлі ісік ауруларының қаупін азайтумен байланысты болды. Шикі жапырақты жасылдар сонымен қатар маңызды В дәрумені фолийіне, сонымен қатар көзді қорғауға көмектесетін лютеин мен зеаксантинге, каротиноидтарға бай. Майда еритін фитохимиялық заттар, мысалы, салат, шпинат, қырыққабат, жасыл және қыша жасыл салаттарда кездесетін каротиноидтар да денеде антиоксиданттық және қабынуға қарсы әсерге ие.
жаңғақтар
Жаңғақтар – төмен гликемиялық тағам және пайдалы майлардың, өсімдік ақуызының, талшықтардың, антиоксиданттардың, фитостеролдардың және минералдардың тамаша көзі, бұл сонымен қатар бүкіл тағамның гликемиялық жүктемесін азайтуға көмектеседі, бұл оларды қант диабетіне қарсы маңызды құрамдас бөлікке айналдырады. диета. Олардың калориялық тығыздығына қарамастан, жаңғақтарды жеу салмақ жоғалтуға көмектеседі. Жаңғақтар сонымен қатар холестеринді азайтады және жүрек ауруларының қаупін азайтуға көмектеседі.
тұқым
Жаңғақтар сияқты тұқымдар пайдалы майларды, антиоксиданттарды және минералдарды қамтамасыз етеді, дегенмен оларда ақуыз көп және микроэлементтерге бай. Чиа, зығыр және қарасора тұқымдары омега-3 майларына бай. Чиа, зығыр және күнжіт тұқымдары да бай лигнандар немесе сүт безі қатерлі ісігімен күресетін фитоэстрогендер болып табылады. Оның үстіне күнжіт дәндері кальций мен Е дәруменіне, ал асқабақ тұқымы мырышқа бай.
Жидектер
Жидектер - жүрек денсаулығын жақсартуға көмектесетін антиоксидантқа бай жемістер. Қатысушылар бірнеше апта бойы күн сайын құлпынай немесе көкжидек жеген зерттеу зерттеулері қан қысымының, жалпы және LDL холестеринінің жақсарғанын және тіпті тотығу стрессінің белгілерін хабарлады. Жидектер сонымен қатар ісікке қарсы қасиеттерге ие және қартаюға байланысты когнитивті құлдырауды болдырмауға көмектесетіні көрсетілген.
Анар
Анардағы ең танымал фитохимиялық пуникалагин жемістің антиоксиданттық белсенділігінің жартысынан көбіне жауап береді. Анардың фитохимиялық заттары қатерлі ісікке қарсы, кардиопротекторлық және миға пайдалы қасиеттерге ие. Бір зерттеуде 28 күн бойы күн сайын анар шырынын ішкен егде жастағы адамдар плацебо сусындарын ішкендермен салыстырғанда есте сақтау сынағы бойынша жақсы нәтиже көрсетті.
Атбас бұршақтар
Бұршақтарды және басқа бұршақтарды жеу қандағы қантты теңестіруге, тәбетіңізді азайтуға және тоқ ішек қатерлі ісігінен қорғауға көмектеседі. Бұршақ – қант диабетіне қарсы тағам, ол салмақ жоғалтуға көмектеседі, өйткені олар баяу қорытылады, бұл тамақтан кейін қандағы қанттың жоғарылауын баяулатады және қанықтыру арқылы тағамға құмарлықтың алдын алуға көмектеседі. Аптасына екі рет бұршақ және басқа да бұршақтарды жеу тоқ ішек қатерлі ісігінің қаупін азайтады. Бұршақтарды және қызыл бұршақ, қара бұршақ, ноқат, жасымық және бұршақ сияқты бұршақтарды жеу де басқа қатерлі ісіктерден айтарлықтай қорғайды.
Саңырауқұлақтар
Саңырауқұлақтарды үнемі жеу сүт безі қатерлі ісігінің қаупін азайтумен байланысты. Ақ және Портобелло саңырауқұлақтары сүт безі қатерлі ісігіне қарсы әсіресе пайдалы, өйткені оларда ароматаза ингибиторлары немесе эстроген өндірісін тежейтін қосылыстар бар. Саңырауқұлақтардың қабынуға қарсы әсері бар, сонымен қатар иммундық жасушалардың белсенділігін арттырады, ДНҚ зақымдануының алдын алады, рак клеткаларының өсуін бәсеңдетеді және ангиогенезді тежейді. Саңырауқұлақтарды әрқашан пісіру керек, өйткені шикі саңырауқұлақтарда агаритин деп аталатын потенциалды канцерогенді химиялық зат бар, ол пісіру кезінде айтарлықтай азаяды.
Пияз және сарымсақ
Пияз мен сарымсақ жүрек-қан тамырлары мен иммундық жүйеге пайдалы әсер етеді, сонымен қатар диабетке және ісікке қарсы әсер етеді. Бұл сонымен қатар асқазан және простата қатерлі ісігінің төмен қаупімен байланысты. Пияз мен сарымсақ канцерогендерді детоксикациялау, ісік жасушаларының өсуін азайту және ангиогенезді тежеу арқылы қатерлі ісіктердің дамуын болдырмауға көмектесетін күкірт органикалық қосылыстарымен танымал. Пияз бен сарымсақ сонымен қатар ісіктің алдын алуға көмектесетін қабынуға қарсы әсері бар денсаулықты жақсартатын флавоноидты антиоксиданттардың жоғары концентрациясына ие.
қызанақ
Қызанақ ликопен, С және Е дәрумендері, бета-каротин және флавонол антиоксиданттары сияқты әртүрлі қоректік заттарға бай. Ликопен простата обырынан, терінің ультракүлгін сәулеленуінен қорғауға көмектеседі және? жүрек - қан тамырлары ауруы. Ликопен қызанақ пісірілген кезде жақсы сіңеді. Бір кесе қызанақ соусында шикі, туралған қызанақтың бір шыныаяқынан шамамен 10 есе көп ликопен бар. Сондай-ақ, ликопен сияқты каротиноидтар пайдалы майлармен бірге жақсы сіңетінін есте сақтаңыз, сондықтан қосымша қоректік артықшылықтар үшін жаңғақ қосылған салатта немесе жаңғақ негізіндегі таңғышта қызанақпен ләззат алыңыз.
Біз тұтынатын тағамдардың денсаулығымызға пайдалы немесе зиянды болуы мүмкін. Нашар тамақтану денсаулықтың әртүрлі мәселелерін, соның ішінде семіздік, жүрек-қан тамырлары аурулары және 2 типті қант диабетін тудыруы мүмкін. Сонымен қатар, дұрыс тамақтану сізді қуаттандырады, денсаулыққа қатысты мәселелер қаупін азайтады, сонымен қатар салауатты салмақты сақтауға және реттеуге көмектеседі. Ұзақ өмір сүруді қаласаңыз, денеңізді жақсы тағамдармен толтыруыңыз керек. Жақсы тағамдар сонымен қатар әртүрлі денсаулық мәселелерімен, соның ішінде буын ауруы мен артритпен байланысты қабынуды азайтуға көмектеседі. Хиропрактиктер сияқты денсаулық сақтау мамандары денсаулық пен денсаулықты нығайтуға көмектесу үшін диета және өмір салты бойынша кеңес ұсына алады. Келесі мақалада біз ұзақ өмір сүруге көмектесетін бірнеше жақсы тағамдарды тізімдейміз. – Доктор Алекс Хименес DC, CCST Insight
Қызғылт сары шырын
Серверлер: 1 Пісіру уақыты: 5-10 минут
1 грейпфрут, қабығы аршылған және туралған
1 алма, жуылған және туралған
1 тұтас қызылша, егер сізде бар болса, оларды жуыңыз және тураңыз
1 дюймдік зімбірдің тұтқасы, шайылған, қабығы және туралған
Барлық ингредиенттерді жоғары сапалы шырын сыққыштан алыңыз. Жақсы бірден қызмет етеді.
Бір сәбіз сізге күнделікті А дәрумені қабылдауының барлығын береді
Иә, бір ғана қайнатылған 80 г (2 унция) сәбізді жеу денеңізге 1,480 микрограмм (мкг) А витаминін (тері жасушаларының жаңаруы үшін қажет) өндіруге жеткілікті бета-каротин береді. Бұл Құрама Штаттардағы А дәруменінің ұсынылатын тәуліктік мөлшерінен көп, яғни шамамен 900 мкг. Пісірілген сәбізді жеген дұрыс, өйткені бұл жасуша қабырғаларын жұмсартады және бета-каротиннің көбірек сіңірілуіне мүмкіндік береді. Сіздің диетаңызға пайдалы тағамдарды қосу - жалпы денсаулықты жақсартудың тамаша тәсілі.
Біздің ақпарат шеңберіміз хиропрактикалық, тірек-қимыл аппараты, физикалық дәрі-дәрмектермен, сауықтыру және денсаулыққа қатысты сезімтал мәселелермен және / немесе функционалды медицина туралы мақалалармен, тақырыптармен және пікірталастармен шектеледі. Біз тірек-қимыл аппаратының жарақаттары немесе бұзылулары кезінде емдеуді қолдау және қолдау үшін функционалды денсаулық пен сауықтыру хаттамаларын қолданамыз. Біздің жазбаларымыз, тақырыптарымыз, тақырыптарымыз бен түсініктеріміз клиникалық мәселелерді, тікелей немесе жанама түрде біздің клиникалық практикамен байланысты және қолдайтын тақырыптарды, мәселелерді және тақырыптарды қамтиды. * Біздің кеңсе қолдайтын дәйексөздер келтіруге негізделген талпыныстар жасады және тиісті зерттеу жұмысын анықтады немесе біздің жазбаларымызды қолдайтын зерттеулер. Сондай-ақ, біз қосымша зерттеулердің көшірмелерін кеңестің және халықтың сұрауы бойынша қол жетімді етіп жасаймыз. Біз оның белгілі бір күтім жоспарына немесе емдеу хаттамасына қалай көмектесуі мүмкін екендігі туралы қосымша түсініктеме талап ететін мәселелерді қарастыратынымызды түсінеміз; сондықтан жоғарыда аталған тақырыпты одан әрі талқылау үшін доктор Алекс Хименестен немесе бізбен байланысыңыз 915-850-0900. Лицензиясы бар провайдерлер (лер) Техаста * және Нью-Мексико *
Доктор Алекс Хименес DC, CCST басқарды
Әдебиеттер тізімі:
Джоэль Фюрман, медицина ғылымдарының докторы. �Ұзақ өмір сүру және сау болу үшін жеуге болатын ең жақсы 10 тағам.� Велвелл денсаулығы, 6 маусым 2020 жыл, www.verywellhealth.com/best-foods-for-longevity-4005852.
Дауден, Анжела. Кофе - бұл жеміс-жидек және басқа да керемет азық-түлік фактілері. MSN өмір салты, 4 маусым 2020 жыл, www.msn.com/en-us/foodanddrink/did-you-know/coffee-is-a-fruit-and-other-unbelievably-true-food-facts/ss-BB152Q5q?li=BBnb7Kz&ocid =mailsignout#image=24.
Егер сіз осындай жағдайлардың кез-келгеніне тап болсаңыз, онда сіздің коллаген пептидтеріңіз төмен болуы мүмкін.
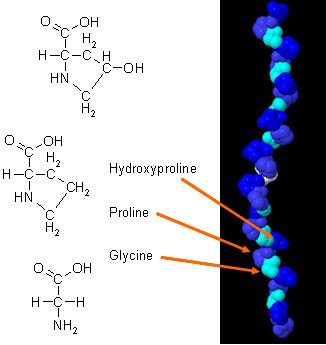
Онда бар жаңа зерттеулер болды күнделікті жаттығулармен үйлескен кезде коллагеннің дене құрамын қалай жақсартуға болатындығы туралы. Денедегі коллаген ағзаның анатомиясында маңызды рөл атқаратын ерекше амин қышқылдарының құрамына ие. Коллаген ақуызы глицин, пролин және гидроксипролиннің шоғырланған көзі болып табылады, және оны барлық диеталық ақуыздармен салыстырған кезде ол коллагенді құрылымдық ақуыз ретінде ықтимал практикалық таңдау жасайды.
In а 2015 зерттеу, зерттеушілер белсенді коллаген қоспаларының белсенді еркектердегі дене құрамын жақсарта алатындығын көрсетті. Нәтижелер әрбір ер адамның салмақ жаттығуларына аптасына кемінде үш рет қатысатындығын және денсаулықты барынша жақсарту үшін кем дегенде 15 грамм коллаген пептидтерін қосу керектігін көрсетеді. Тесттің күші сынақ, биоимпеданс анализі (BIA) және бұлшықет биопсиясы болып табылады. Бұл сынақтар еркектер коллаген қоспаларын қабылдағаннан кейін жақсы жұмыс істейтініне көз жеткізеді және нәтижелер олардың дене салмағының майсыз дене салмағының жоғарылағанын көрсетеді. Тағы бір зерттеу егде жастағы адамдармен, сондай-ақ саркопениямен ауыратын адамдарға бұлшықет массасын және бұлшықет күшін арттыратын қарсылық жаттығуларымен біріктірілген кезде коллаген протеинін қалай қосуды көрсетті.
Коллагеннің пайдалы қасиеттері
Сонда бар көптеген пайдалы қасиеттері бар бұл коллаген қоспалары тұтынылған кезде ағзаға бере алады. Гидролиздендірілген коллаген мен желатин бар, олар адамның тері құрылымын жақсартуға көмектеседі. Коллагендік қоспалар туралы көптеген зерттеулер болмаса да, дененің аумақтары үшін керемет уәделер бар. Олар:
Бұлшықеттер массасы: Коллаген қоспалары күш жаттығуларымен бірге дененің бұлшықет массасы мен күшін арттырады.
артрит: Коллаген қоспалары остеоартритпен ауыратын адамдарға көмектеседі. Зерттеулер көрсетті адамдар остеоартритте коллаген қоспаларын қабылдаған кезде, олар ауырсынудың жаппай төмендегенін анықтады.
Тері серпімділігі: In а 2014 зерттеу, коллаген қоспаларын қабылдаған және терінің икемділігі жақсарған әйелдер туралы айтылды. Коллагенді жергілікті терапия кезінде де қолдануға болады, ол терінің сыртқы көрінісін жақсартуға көмектеседі, ол әжімдер мен әжімдерді азайтады.
Коллаген қоспалары дененің белгілі бір аймақтарына пайдалы қасиеттер беріп қана қоймайды, сонымен қатар коллагеннің төрт негізгі түрі бар және олардың адам ағзасындағы рөлі, сондай-ақ атқаратын қызметтері:
1 теріңіз: 1 типті коллаген ағзадағы коллагеннің 90% құрайды және терінің, сүйектердің, дәнекер тіндерінің және денеде орналасқан тістердің құрылымын қамтамасыз ететін тығыз оралған талшықтардан тұрады.
2 теріңіз: 2 типті коллаген серпімді шеміршекте орналасқан бос оралған талшықтардан құралған, бұл денеде буындарды басуға көмектеседі.
3 теріңіз3 типті коллаген дененің дұрыс жұмыс істейтініне көз жеткізетін бұлшықеттердің, мүшелер мен артериялардың құрылымын қолдауға көмектеседі.
4 теріңіз4 типті коллаген әр адамның терісінің қабаттарында кездеседі және ағзадағы сүзілуге көмектеседі.
Коллагеннің осы төрт түрі ағзада болғандықтан, уақыт өткен сайын коллагеннің табиғи түрде азаюы мүмкін екенін білу керек, өйткені денеде коллагеннің сапасы төменірек болады. Коллагеннің төмендеуінің көрінетін белгілерінің бірі - бұл адам ағзасындағы тері әлсіз болып, серпіліп, қартаюға байланысты шеміршек әлсіреуі.
Коллагенді зақымдауы мүмкін факторлар
Коллаген жасына қарай табиғи түрде азаюына қарамастан, көптеген факторлар теріге зиянды коллагенді жоя алады. Зиянды факторларға мыналар кіруі мүмкін:
Қант және көмірсуларТазартылған қанттар және көмірсулар кедергі келтіруі мүмкін коллагеннің теріні қалпына келтіру қабілеті бар. Демек, ағзадағы қант пен көмірсутектерді тұтынуды азайту арқылы ол қан тамырлары, бүйрек және тері тіндерінің дисфункциясының әсерін азайтады.
Күн сәулесі: Күн жеткілікті болса да, адамға күнді рахаттана көруге көмектеседі, алайда ұзақ уақыт бойы күн сәулесінде болады зақым келтіруі мүмкін теріге түсіп, коллаген пептидтерін бұзады. Күннің шамадан тыс әсер етуі терінің фото жасына әкеліп, денеде тотығу стрессін тудыруы мүмкін.
Шылым шегу: Адам темекі шегетін кезде, ол мүмкін коллаген өндірісін азайту денеде ерте әжімдер пайда болады, егер дене жараланса, сауығу процесі баяу жүреді және денеде ауруға әкелуі мүмкін.
Автоиммунды аурулар: Кейбір аутоиммундық аурулар лупус тәрізді коллаген өндірісін зақымдауы мүмкін.
қорытынды
Коллаген ағза үшін өте маңызды, өйткені терінің жұмсақ және қатты болуына көмектеседі. Әрине, адам қартайған сайын азаяды, сондықтан коллаген қоспаларын қабылдау дененің дұрыс жұмыс істей алатындығына көз жеткізеді. Денеге зиянды факторлар әсер етсе, олар коллаген өндірісін тоқтата алады немесе тіпті зақымдауы мүмкін және ерте әжімдердің пайда болу процесін тездетеді, бұл адамды олардан үлкенірек етеді. Кейбір өнімдер дененің жасушалық белсенділігіне, тұрақтылықты, биожетімділігі мен асқорытудың жайлылығын қамтамасыз ете алады.
Біздің ақпаратымыздың шеңбері хиропрактика, тірек-қимыл және жүйке денсаулығына қатысты мәселелер немесе функционалды медицина туралы мақалалармен, тақырыптармен және пікірталастармен шектеледі. Біз тірек-қимыл аппаратының жарақаттары мен бұзылуларын емдеу үшін денсаулық сақтаудың функционалды хаттамаларын қолданамыз. Біздің кеңсе тірек дәйексөздерді ұсынуға ақылға қонымды әрекет жасады және біздің зерттеуімізді немесе біздің посттарды қолдайтын зерттеулерді анықтады. Біз сондай-ақ қолдауға ие болған зерттеулердің көшірмелерін басқармаға немесе сұраныс бойынша көпшілікке ұсынамыз. Жоғарыда аталған тақырыпты одан әрі талқылау үшін доктор Алекс Хименеске хабарласыңыз немесе бізбен байланысыңыз 915-850-0900.
Әдебиеттер тізімі:
Бош, Рикардо және т.б. Фотохимия және тері фотокарциногенезінің механизмдері және фитохимиялық заттармен фотопротективті стратегиялар. Антиоксиданттар (Базель, Швейцария), MDPI, 26 наурыз 2015 ж., Www.ncbi.nlm.nih.gov/pmc/articles/PMC4665475/.
Дэнби, Ф. Уильям. Тамақтану және қартаю терісі: қант және гликация.ation Дерматологиядағы клиникалар, АҚШ ұлттық медицина кітапханасы, 2010, www.ncbi.nlm.nih.gov/pubmed/20620757.
Дженнингс, Керри-Анн. Коллаген - бұл не және ол не үшін пайдалы? Денсаулық жолы, 9 қыркүйек 2016 ж., Www.healthline.com/nutrition/collagen.
Юргелевич, Майкл. Жаңа зерттеу коллаген пептидтерінің дене жаттығуларымен үйлесетін құрамын жақсартудың артықшылықтарын көрсетеді. Денсаулыққа арналған дизайн, 31 мамыр 2019, blog.designsforhealth.com/node/1031.
Кнутинен, А және т.б. Шылым шегу коллаген синтезіне және адамның терідегі жасушадан тыс матрицалық айналымына әсер етеді. Британдық дерматология журналы, АҚШ Ұлттық медицина кітапханасы, сәуір, 2002, www.ncbi.nlm.nih.gov/pubmed/11966688.
Proksch, E және басқалар. Арнайы коллаген пептидтерінің ауызша қосылуы адамның тері физиологиясына пайдалы әсер етеді: қос соқыр, плацебо бақыланатын зерттеу. Тері фармакологиясы және физиологиясы, АҚШ ұлттық медицина кітапханасы, 2014, www.ncbi.nlm.nih.gov/pubmed/23949208.
Шаусс, Александр Г және т.б. Остеоартритке байланысты белгілерді жақсартуға арналған төмен молекулалық салмағы бар гидролизденген тауық еті шеміршек сіріндісі, биоцелл коллагенінің романының әсері: кездейсоқ, қос соқыр, плацебо бақыланатын сынақ. Ауыл шаруашылығы және азық-түлік химия журналы, АҚШ Ұлттық медицина кітапханасы, 25 сәуір, 2012, www.ncbi.nlm.nih.gov/pubmed/22486722.
Здзиеблик, Дениз және т.б. Коллаген пептидінің қосылуы қарсылық жаттығуларымен бірге ағзаның құрамын жақсартады және егде жастағы саркопениялық ерлердегі бұлшықет күшін жоғарылатады: кездейсоқ бақыланатын сынақ. Британдық тамақтану журналы, Кембридж Университетінің Баспасы, 28 қазан 2015 ж., Www.ncbi.nlm.nih.gov/pmc/articles/PMC4594048/.
Ұлттық денсаулық сақтау ғылымдары университеті болашақ ұрпаққа білімді қалай беретіні туралы адамдарға ақпарат бере отырып, университет функционалды медицина бойынша көптеген медициналық мамандықтарды ұсынады.
Сізге целиак ауруы, тітіркенген ішек синдромы, дивертикулоз/дивертикулит немесе ағып жатқан ішек синдромы диагнозы қойылған сияқты ма?
Шамадан тыс кекіру, кекіру немесе ісіну?
Белгілі бір пробиотиктер немесе табиғи қоспалардан кейін қалыптан тыс созылу?
Қоректік заттардың сіңірілуіне күдік?
Релаксациямен ас қорыту проблемалары басыла ма?
Егер сізде осы жағдайлардың кез келгені болса, онда сізде ішек проблемалары болуы мүмкін және 4R бағдарламасын қолданып көру керек болуы мүмкін.
Тағамға сезімталдық, ревматоидты артрит және алаңдаушылық асқазан-ішек жолдарының өткізгіштігінің бұзылуымен байланысты болды. Бұл әртүрлі жағдайлар ас қорыту жолына әсер етуі мүмкін көптеген факторлардан туындауы мүмкін. Егер емделмеген болса, бұл ішектің өткізгіштігінің тосқауылының дисфункциясының нәтижесі болуы мүмкін, қабынуды тудыруы және ішек дамуы мүмкін ауыр денсаулық жағдайлары болуы мүмкін. 4R бағдарламасы ағзадағы сау ішекті қалпына келтіру үшін қолданылады және төрт қадамды қамтиды. Олар: алып тастау, ауыстыру, қайта егу және жөндеу.
Ішектің өткізгіштігі
Ішектің өткізгіштігі денені қорғауға көмектеседі және зиянды бактериялардың ішекке түспеуін қамтамасыз етеді. Ол денені одан қорғайды ықтимал экологиялық факторлар Бұл зиянды болуы мүмкін және ас қорыту жолдары арқылы енеді. Бұл ас қорыту жолдарына зиян келтіретін токсиндер, патогендік микроорганизмдер және басқа антигендер болуы мүмкін. Ішектің кілегей қабаты тығыз түйіспелермен бөлінген эпителий жасушаларының қабатынан тұрады. Сау ішекте, тығыз түйісу заттардың ішек тосқауылына селективті түрде енуіне және өтуіне және зиянды факторлардың сіңірілуіне жол бермеу арқылы ішектің өткізгіштігін реттейді.
Қоршаған ортаның кейбір факторлары тығыз түйіспеге зақым келтіруі мүмкін, нәтижесінде ол ішектің өткізгіштігін жоғарылатуы мүмкін, бұл ішектің гиперөткізгіштігін немесе ағзадағы ағып кетуді тудырады. Қаныққан майлар мен алкогольдің шамадан тыс мөлшері, қоректік заттардың жетіспеушілігі, созылмалы стресс және жұқпалы аурулар сияқты әсер ететін факторлар ішектің өткізгіштігін арттыруы мүмкін.
Ішек өткізгіштігінің жоғарылауымен ішекте ол антигендердің ішектің шырышты қабығынан өтіп, қанға енуіне мүмкіндік береді, бұл иммундық жауап пен дененің қабынуын тудырады. Ішектің гиперөткізгіштігімен байланысты асқазан-ішек жолдарының белгілі бір жағдайлары бар және олар емделмеген жағдайда денеге зиян келтіруі мүмкін кейбір аутоиммундық жағдайларды тудыруы мүмкін.
4Rs бағдарламасы
4Rs - бұл денсаулық сақтау мамандары пациенттеріне ас қорытуды бұзу мәселелерін шешкен кезде қолдануға кеңес беретін және ішектің жазылуына көмектесетін бағдарлама.
Мәселені жою
4Rs бағдарламасындағы бірінші қадам ішек өткізгіштігінің жоғарылауымен байланысты зиянды патогенді және қабыну триггерлерін жою болып табылады. Стресс және созылмалы алкогольді тұтыну сияқты триггерлер адамның денесіне көп зиян келтіруі мүмкін. Осы зиянды факторларды организмнен мақсатты түрде емдеу оны дәрі-дәрмекпен, антибиотиктермен, қоспалармен емдеу болып табылады және диетадан қабынуды тудыратын тағамдарды алып тастау ұсынылады, соның ішінде:
- Алкоголь
- Глютен
– Тағамдық қоспалар
– Крахмалдар
- Кейбір май қышқылдары
– Адам сезімтал болатын кейбір тағамдар
Қоректік заттарды ауыстыру
4Rs бағдарламасының екінші қадамы қабыну арқылы ішек проблемаларын тудыратын қоректік заттарды ауыстыру болып табылады. Кейбір қоректік заттар ас қорыту трактінің қолдау көрсетілетініне көз жеткізе отырып, ішектегі қабынуды азайтуға көмектеседі. Қабынуға қарсы кейбір қоректік тағамдар бар. Оларға мыналар жатады:
– Талшықтылығы жоғары тағамдар
– Омега-3
- Зәйтүн майы
- Саңырауқұлақтар
- Қабынуға қарсы шөптер
Салауатты ішекті жақсарту үшін қоректік заттарды сіңіру және сіңіру арқылы ас қорыту функциясын қолдау үшін белгілі бір қоспалар бар. Асқорыту ферменттері ішектегі майларды, ақуыздарды және көмірсуларды ыдыратуға көмектеседі. Бұл ас қорыту жолы бұзылған, тағамға төзімсіздігі бар немесе целиак ауруы бар адамдарға пайдалы болады. Өт қышқылы қоспалары сияқты қоспалар липидтерді біріктіру арқылы қоректік заттардың сіңуіне көмектесуі мүмкін. Зерттеулер мәлімдеген өт қышқылдары бауырды, өт қабын және өт жолын емдеу үшін қолданылғанын, сонымен бірге бариатриялық хирургиядан кейін өт тастарының пайда болуын болдырмайды.
Ішек қайта егілді
Үшінші қадам - ішек микробын пайдалы бактериялармен қайта егуге арналған 4rs бағдарламасының сау ішек жұмысын ынталандыру. Зерттеулер көрсетілген пробиотикалық қоспалар пайдалы бактерияларды қалпына келтіру арқылы ішекті жақсарту үшін қолданылған. Бұл қоспалардың көмегімен олар ағзаға қабынуға қарсы заттарды бөліп шығару арқылы ішектің жақсаруын қамтамасыз етеді, иммундық жүйені қолдауға, дененің микробтық құрамын өзгертуге және ішек жүйесіндегі ішектің өткізгіштігін төмендетуге көмектеседі.
содан бері пробиотиктер кездеседі ашытылған тағамдарда және өтпелі деп саналады, өйткені олар асқазан-ішек жолында тұрақты емес және пайдалы. Бір таңқаларлығы, олар витаминдер мен микробқа қарсы қосылыстар шығару арқылы ішекке әсер ету арқылы адам денсаулығына әсер етеді, осылайша әртүрлілік пен ішек жұмысын қамтамасыз етеді.
Ішекті жөндеу
4Rs бағдарламасының соңғы қадамы ішекті жөндеу болып табылады. Бұл қадам арнайы қоректік заттар мен шөптермен ішектің ішек қабығын қалпына келтіруді қамтиды. Бұл шөптер мен қоспалар ішектің өткізгіштігін және ағзадағы қабынуды азайтуға көмектеседі. Бұл шөптер мен қоспалардың кейбірі мыналарды қамтиды:
– Алоэ вера
– Хиос мастикасы
– DGL (деглицирризизацияланған мия)
– Зефир тамыры
- L-глутамин
– Омега-3
� Полифенолдар
- D дәрумені
- Мырыш
қорытынды
Өйткені көптеген факторлар ас қорыту жүйесіне зиянды әсер етуі мүмкін және бірнеше денсаулық жағдайына себепші болуы мүмкін. 4Rs бағдарламасының негізгі мақсаты - ішекке зиян келтіретін және қабынуды азайтатын және ішек өткізгіштігін арттыратын осы факторларды азайту. Науқас 4R қамтамасыз ететін пайдалы факторлармен таныстырылған кезде, бұл сау, емделген ішекке әкелуі мүмкін. Кейбір өнімдер ішектерді қолдау, қант алмасуын жақсарту және ішектерді қолдауға арналған амин қышқылдарына бағытталған асқазан-ішек жүйесін қолдауға көмектесу үшін осында.
Біздің ақпаратымыздың шеңбері хиропрактика, тірек-қимыл және жүйке денсаулығына қатысты мәселелер немесе функционалды медицина туралы мақалалармен, тақырыптармен және пікірталастармен шектеледі. Біз тірек-қимыл аппаратының жарақаттары мен бұзылуларын емдеу үшін денсаулық сақтаудың функционалды хаттамаларын қолданамыз. Біздің кеңсе тірек дәйексөздерді ұсынуға ақылға қонымды әрекет жасады және біздің зерттеуімізді немесе біздің посттарды қолдайтын зерттеулерді анықтады. Біз сондай-ақ қолдауға ие болған зерттеулердің көшірмелерін басқармаға немесе сұраныс бойынша көпшілікке ұсынамыз. Жоғарыда аталған тақырыпты одан әрі талқылау үшін доктор Алекс Хименеске хабарласыңыз немесе бізбен байланысыңыз 915-850-0900.
Әдебиеттер тізімі:
Де Сантис, Стефания және т.б. �Ішек тосқауылының модуляциясының қоректік кілттері.� Иммунологиядағы шекаралар, Frontiers Media SA, 7 желтоқсан, 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4670985/.
Ианиро, Джанлука және т.б. �Асқазан-ішек аурулары кезіндегі ас қорыту ферменттерін толықтыру.� Қазіргі дәрілік зат алмасу, Bentham Science Publishers, 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC4923703/.
Му, Цинхуи және т.б. �Ішектің ағуы аутоиммундық аурулардың қауіпті сигналы ретінде.� шекара, Шекаралар, 5 мамыр 2017 жыл, www.frontiersin.org/articles/10.3389/fimmu.2017.00598/full.
Резак, Шеннон және т.б. �Ашытылған тағамдар тірі ағзалардың диеталық көзі ретінде.� Микробиологиядағы шекаралар, Frontiers Media SA, 24 тамыз 2018 жыл, www.ncbi.nlm.nih.gov/pmc/articles/PMC6117398/.
Сандер, Гай Р., т.б. �Глиадин арқылы ішек тосқауылының функциясын жылдам бұзу апикальды қосылыс белоктарының экспрессиясының өзгеруін қамтиды.� FEBS баспасөзі, John Wiley & Sons, Ltd, 8 тамыз 2005 ж., febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2005.07.066.
Сартор, Р.Балфур. �Ішектің қабыну ауруларындағы ішек микрофлорасын емдік манипуляциялау: антибиотиктер, пробиотиктер және пребиотиктер.� Гастроэнтерология, АҚШ Ұлттық медицина кітапханасы, мамыр 2004, www.ncbi.nlm.nih.gov/pubmed/15168372.
Созылмалы ауырсыну Америка Құрама Штаттарындағы көптеген адамдарға әсер ететін жалпы денсаулық мәселесі болып табылады. Фибромиалгия және миофассиялық ауырсыну синдромы сияқты бірнеше медициналық жағдайлар созылмалы ауырсынуды тудыруы мүмкін, бірақ ол денсаулықтың басқа да мәселелеріне байланысты дамуы мүмкін. Зерттеулер кең таралған қабыну созылмалы ауырсынудың негізгі себебі екенін анықтады. Қабыну - жарақаттан, аурудан немесе инфекциядан табиғи қорғаныс механизмі. Бірақ, егер қабыну процесі тым ұзақ уақытқа созылса, ол проблемаға айналуы мүмкін.
Қабыну иммундық жүйеге зақымдалған тіндерді емдеуге және қалпына келтіруге, сондай-ақ өзін бактериялар мен вирустардан қорғауға сигнал береді. Жоғарыда айтылғандай, созылмалы қабыну әртүрлі денсаулық мәселелерін, соның ішінде созылмалы ауырсыну белгілерін тудыруы мүмкін. Салауатты өмір салтын өзгерту созылмалы ауырсынуды жеңуге көмектеседі, бірақ алдымен созылмалы ауырсынудың жалпы себептерін түсінейік.
Жедел қабыну дегеніміз не?
Жедел қабыну, мысалы, жарақаттан кейін немесе тамақ ауруы сияқты қарапайым нәрседен кейін пайда болады. Бұл жағымсыз әсерлері бар табиғи жауап, яғни ол денсаулық мәселесі табылған аймақта жергілікті деңгейде жұмыс істейді. Ұлттық медицина кітапханасы мәлімдегендей, жедел қабынудың жалпы белгілеріне ісіну, қызару, жылу, ауырсыну және функцияның жоғалуы жатады. Жедел қабыну дамыған кезде қан тамырлары кеңейіп, қан ағымының жоғарылауына әкеледі, ал зақымдалған аймақтағы лейкоциттер қалпына келтіруге ықпал етеді.
Қатты қабыну кезінде зақымдалған тіннен цитокиндер деп аталатын қосылыстар бөлінеді. Цитокиндер адам ағзасының иммундық жасушаларына, сондай-ақ денсаулық мәселесін жөндеу үшін гормондар мен көптеген қоректік заттарға әкелетін «төтенше сигналдар» ретінде әрекет етеді. Сонымен қатар, простагландиндер деп аталатын гормонға ұқсас заттар зақымдалған тіндерді емдеу үшін қан ұйығыштарын тудырады және олар қабыну процедурасының бөлігі ретінде қызба мен ауырсынуды тудыруы мүмкін. Зақымдану немесе жарақат қалпына келтірілгенде, қабыну басылады.
Созылмалы қабыну дегеніміз не?
Жедел қабынудан айырмашылығы, созылмалы қабыну ұзақ мерзімді әсер етеді. Созылмалы қабыну, сонымен қатар тұрақты қабыну деп аталады, қан мен жасуша тіндерінде орналасқан иммундық жүйе маркерлерінің жоғарылауымен көрсетілгендей, бүкіл адам ағзасында қабынудың төмен деңгейін тудырады. Созылмалы қабыну әртүрлі аурулар мен жағдайлардың дамуын тудыруы мүмкін. Қабынудың жоғары деңгейі кейде жарақат, ауру немесе инфекция болмаса да, иммундық жүйенің реакциясын тудыруы мүмкін.
Нәтижесінде адам ағзасының иммундық жүйесі сау жасушаларға, тіндерге немесе мүшелерге шабуыл жасай бастайды. Зерттеушілер әлі күнге дейін адам ағзасындағы созылмалы қабынудың салдарын және осы табиғи қорғаныс процесіне қатысатын механизмдерді түсінуге тырысуда. Мысалы, созылмалы қабыну жүрек ауруы және инсульт сияқты әртүрлі денсаулық мәселелерімен байланысты болды.
Бір теория қабыну қан тамырларында қалған кезде, ол бляшканың жиналуын ынталандыруы мүмкін деп болжайды. Американдық жүрек қауымдастығының немесе AHA-ның мәліметтері бойынша, егер иммундық жүйе бляшканы бөтен басқыншы ретінде анықтаса, лейкоциттер артериялар арқылы ағып жатқан қандағы бляшкаларды жабуға тырысуы мүмкін. Бұл қан ұйығышын тудыруы мүмкін, ол жүрекке немесе миға қан ағынын бөгеп, оның тұрақсыз болуына және жарылуына әкелуі мүмкін. Қатерлі ісік - созылмалы қабынумен байланысты денсаулықтың тағы бір мәселесі. Сонымен қатар, Ұлттық онкологиялық институттың мәліметтері бойынша, ДНҚ зақымдануы созылмалы қабынудан да туындауы мүмкін.
Тұрақты, төмен дәрежелі қабынудың жиі белгілері болмайды, бірақ медицина мамандары С-реактивті ақуызды немесе қандағы қабыну маркері липой қышқылы деп аталатын CRP-ны тексере алады. CRP деңгейінің жоғарылауы жүрек-қан тамырлары ауруларының қаупінің жоғарылауымен байланысты. CRP деңгейінің жоғарылауы қызыл жегі немесе ревматоидты артрит сияқты созылмалы ауруларда болуы мүмкін.
Фибромиалгия сияқты басқа созылмалы жағдайларда жүйке жүйесі арнайы ынталандыруға шамадан тыс әрекет етеді, бірақ бұл созылмалы ауырсыну белгілерін тудыратын қабыну. Субъективті түрде жүйке жүйесінен туындаған созылмалы ауырсыну мен кең таралған қабынудан туындаған созылмалы ауырсыну арасындағы айырмашылықты айту мүмкін емес. Қан ағымындағы анықтамаларды іздеуден басқа, адамның тамақтануы, өмір салты және қоршаған ортаға әсер етуі созылмалы қабынуды тудыруы мүмкін.
Қабыну иммундық жүйенің жарақаттан, аурудан немесе инфекциядан табиғи қорғаныс механизмі болып табылады. Бұл қабыну реакциясы тіндерді емдеуге және қалпына келтіруге көмектессе, созылмалы, кең таралған қабыну әртүрлі денсаулық мәселелерін, соның ішінде созылмалы ауырсыну белгілерін тудыруы мүмкін. Теңдестірілген тамақтану, соның ішінде әртүрлі диеталар мен ораза, қабынуды азайтуға көмектеседі. Ораза, сондай-ақ калориялық шектеу ретінде белгілі, жасуша апоптозы мен митохондрияның қалпына келуіне ықпал етеді. Ұзақ өмір сүруге арналған диета жоспарының бөлігі болып табылатын ораза ұстауға еліктеу диетасы - дәстүрлі оразаның артықшылықтарын сезіну үшін адам ағзасын ораза күйіне «алдайтын» диеталық бағдарлама. Осы мақалада сипатталған диеталардың кез келгенін орындамас бұрын, дәрігермен кеңесуді ұмытпаңыз.
Доктор Алекс Хименес DC, CCST Insight
Тамақтану, диеталар, ораза және созылмалы ауырсыну
Қабынуға қарсы диеталар негізінен жаңа піскен жемістер мен көкөністерді, балық пен майларды тұтынудан тұрады. Мысалы, Жерорта теңізі диетасының жоспары қабынуға қарсы диета болып табылады, ол қалыпты мөлшерде жаңғақтарды жеуге, өте аз ет жеуге және шарап ішуге ықпал етеді. Омега-3 май қышқылдары сияқты қабынуға қарсы тағамдық бөліктер адам ағзасын зиянды әсерлерден қорғайды. daмаз қабыну нәтижесінде пайда болады.
Қабынуға қарсы диета сонымен қатар қабынуды тудыруы мүмкін тағамдардан аулақ болуды қамтиды. Ет сияқты транс және қаныққан майлары көп тұтынылатын тағамдардың мөлшерін азайту өте қолайлы. Сонымен қатар, қабынуға қарсы диета тазартылған көмірсулар мен нан мен күріш сияқты тағамдарды тұтынуды шектейді. Олар сондай-ақ маргарин мен күнбағыс, мақсары сияқты омега-6 май қышқылдары бар майларды пайдалануды қысқартуға ықпал етеді. және жүгері майлары.
Ораза ұстау немесе калорияны шектеу тотығу стрессін төмендететін және әртүрлі ағзалардағы қартаю механизмдерін бәсеңдететіні бұрыннан белгілі. Оразаның әсерлері бағдарламаланған жасуша өлімін немесе апоптозды, транскрипцияны, мобильді энергия тиімділігін, митохондриялық биогенезді, антиоксиданттық механизмдерді және циркадиялық ырғақты қамтиды. Ораза сонымен қатар митохондриялардағы гендер митохондрияның қалпына келуіне ықпал ететін апоптозға ұшырау үшін ынталандырылатын митохондрия деп аталатын митохондриялық аутофагияға ықпал етеді.
Үзіліссіз ораза қабынумен күресуге, ас қорытуды жақсартуға және өмір сүру ұзақтығын арттыруға көмектеседі. Адам ағзасы ұзақ уақыт бойы тамақсыз өмір сүре алатындай етіп жасалған. Зерттеулер үзіліссіз аштық ішек микробиотасының жалпы құрамында оң өзгерістерге әкелуі мүмкін екенін көрсетті. Сонымен қатар, үзіліссіз ораза иммундық жүйенің реакциясын арттыра отырып, инсулинге төзімділікті төмендетеді. Соңында, үзіліссіз ораза қабыну ауруларына қатысатын иммундық жүйенің бір бөлігін блоктайтын, сондай-ақ цитокиндер мен С-реактивті ақуыз сияқты қабыну маркерлерінің өндірісін айтарлықтай төмендететін ?-гидроксибутират деп аталатын заттың өндірілуіне ықпал етуі мүмкін. , немесе жоғарыда айтылған CRP.
Доктор Вальтер Лонго кітабында ұсынылған Ұзақ өмір сүру диетасының жоспары қабынуды тудыруы мүмкін өңделген тағамдарды тұтынуды болдырмайды, әл-ауқат пен ұзақ өмір сүруге ықпал етеді. Бұл бірегей диеталық бағдарлама, көптеген дәстүрлі диеталардан айырмашылығы, салмақ жоғалтуға ықпал етпейді. Салмақты азайтуды сезінуіңіз мүмкін болса да, бұл бірегей диеталық бағдарламаның басты мақсаты дұрыс тамақтану болып табылады. Ұзақ өмір сүру диетасының жоспары дің жасушаларына негізделген жаңаруды белсендіруге, іштің майын азайтуға және жасқа байланысты сүйектер мен бұлшықеттердің жоғалуын болдырмауға, сондай-ақ жүрек-қан тамырлары ауруларына, Альцгеймер ауруына, қант диабетіне және қатерлі ісікке төзімділікті арттыруға көмектесу үшін көрсетілді.
Аштықты симптомдық диета немесе FMD сіздің денеңіздің тамақтануынсыз дәстүрлі оразаның пайдасын көруге мүмкіндік береді. FMD негізгі айырмашылығы, бірнеше күн немесе тіпті апта бойы барлық тағамдарды толығымен жоюдың орнына сіз тек калорияларды тек айдың бес күнінде ғана шектеп отырасыз. FMD айына бір рет денсаулық пен денсаулықты жақсартуға көмектесе алады.
Кез келген адам FMD-ні жеке-жеке бақылай алады, ал ProLon оразаны имитациялайтын диета әр күн үшін жеке оралған және таңбаланған 5 күндік тағам бағдарламасын ұсынады, ол аусыл ауруына қажет тағамдарды нақты мөлшерде және комбинацияда ұсынады. Тамақтану бағдарламасы жеуге дайын немесе оңай дайындалатын өсімдік тектес тағамдардан, соның ішінде барлар, сорпалар, жеңіл тағамдар, қоспалар, сусын концентраты және шайлардан тұрады. Бастамас бұрын ProLon диетаны имитациялайтын ораза, 5 күндік тамақтану бағдарламасы, немесе жоғарыда сипатталған өмір салтын өзгертулердің кез келгені, сізге қандай созылмалы ауруды емдеу дұрыс екенін білу үшін денсаулық сақтау маманымен сөйлескеніңізге көз жеткізіңіз.
Біздің ақпаратымыздың көлемі широпротикамен, жұлын денсаулығымен және функционалдық медицинадағы мақалалармен, тақырыптармен және пікірталастармен шектеледі. Жоғарыдағы тақырыпты әрі қарай талқылау үшін д-р Алекс Хименеске немесе бізбен хабарласыңыз 915-850-0900 .
Доктор Алекс Хименес мырзаның жетекшілігімен
Қосымша тақырыпты талқылау: Жедел арқа ауруы
Арқа ауруы - мүгедектіктің ең көп таралған себептерінің бірі және бүкіл әлем бойынша жұмыстан босатылған күндер. Кері ауруы дәрігерлік кеңселерге баратын ең көп тараған себептерге байланысты, тек жоғарғы тыныс жолдарының инфекцияларымен шектеледі. Шамамен ХNUMX пайызы халық өмірінде кем дегенде бір рет ауырсынуды сезінеді. Сіздің омыртқаның - бұл жұмсақ маталардан тұратын сүйек, буын, байлам және бұлшықеттерден тұратын күрделі құрылым. Жарақат және / немесе ауырлататын жағдайлар сияқты қатты дискілер, ақыр соңында арқа ауырсыну белгілеріне алып келуі мүмкін. Спорттық жарақаттар немесе автомобиль апатының жарақаттары көбінесе арқадағы ауырсынудың жиі себебі болып табылады, бірақ кейде ең қарапайым қозғалыстың ауыр нәтижесі болуы мүмкін. Бақытымызға орай, широпрактикалық күтім сияқты альтернативті емдеу параметрлері омыртқаның түзетуін және манипуляцияларды қолдану арқылы ауырсынуды жеңілдетуге көмектеседі, ақыр соңында ауруды жеңілдетеді.
XYMOGEN s Эксклюзивті кәсіби формулалар таңдаулы лицензиялық денсаулық сақтау мамандары арқылы қол жетімді. Интернетті сату және XYMOGEN формулаларын дисконттау қатаң тыйым салынады.
Мақтанышпен, Александр Хименес XYMOGEN формулаларын біздің қамқорлығымыздағы науқастарға ғана қол жетімді етеді.
Шұғыл қол жеткізу үшін дәрігерге кеңес беру үшін біздің кеңсемізге хабарласыңыз.
Егер сіз науқас болсаңыз Медициналық және хиропрактикалық жарақаттар клиникасы, сіз қоңырау шалу арқылы XYMOGEN туралы біле аласыз 915-850-0900.
Сізге ыңғайлы болу үшін және шолу XYMOGEN өнімдері төмендегі сілтемемен танысыңыз. *XYMOGEN-Каталог -жүктеу
* Жоғарыда аталған барлық XYMOGEN саясаты қатаң сақталады.
Дұрыс тамақтануды қамтамасыз ету үшін белгілі бір диетаны сақтау кейде стрессті жеуі мүмкін. Табиғи өмір салтын өзгерту - сіздің тағам әдеттеріңізді өзгертудің кілті, бұл сізге ұзақ, сау өмір сүруіңізге көмектеседі. Доктор Валтер Лонго құрған ұзақ өмір сүру диета жоспары жалпы денсаулық пен денсаулыққа қол жеткізу үшін тамақ үлгілерін өзгертуге бағытталған практикалық тағам нұсқаулықтарын таңдау болып табылады.
Ұзақ өмір сүру диетасының жоспары
Төменде келтірілген тамақтану туралы кеңестерді қарап, сіз өзіңіздің ағымдағы диеталарыңыздың жоспарын қайта қарап, дәстүрлі рационның барлық стресстерсіз сау тамақтануды бастай аласыз. Ұзақ мерзімді диета жоспары денсаулыққа қатысты әртүрлі мәселелерді тудыруы және ұзақ өмір сүруге көмектесетін қоректік заттардың тұтынылуын арттыратын өңделген тағамдардың тұтынылуын жояды. Бұл бірегей диеталық бағдарлама шамамен 25 жыл зерттеу жұмыстарының нәтижелерін қарапайым шешім арқылы бөледі, бұл адамдарға тиісті тамақтану арқылы жалпы әл-ауқаттың қаншалықты сезінуіне көмектеседі.
Дегенмен, дәстүрлі диеталардың көпшілігінен айырмашылығы, ұзақ өмір диеталық жоспары салмақ жоғалтуға ықпал етпейді. Сіз салмақты азайтуды сезінсеңіз де, бұл диеталық бағдарламаның маңыздылығы - дұрыс тамақтануға бағытталған. Ұзақ өмірлік диета жоспары сізге жасушалар негізінде жаңаруды белсендіруге, салмақ жоғалтуға және іштің майын азайтуға, жасқа байланысты сүйек пен бұлшықет жоғалтудың алдын алуға, жүрек-қан тамырлары ауруларына, Альцгеймер ауруына, қант диабетіне және қатерлі ісікке қарсы тұруға көмектеседі ұзақ өмір сүруді ұзарту. Төменде біз ұзақ өмір сүру диета жоспарындағы ең көп таралған 8 тамақтану кеңестерін қорытындылаймыз, бұл сіздің өміріңізді ұзақ және сау етуге көмектеседі.
Ұзақ мерзімді диета жоспары - доктор Вальтер Лонгомен жалпы денсаулықты, денсаулықты және ұзақ өмірді дамытуға арналған бірегей диеталық бағдарлама. Адамдар қарапайым өмір салтын өзгерту арқылы тамақтану дағдыларын өзгерте алады және осы диеталық бағдарламаның көптеген артықшылықтарын пайдалана алады. Pescatarian диетасын қадағалау және кейін ProLon ораза ұстаушыларға арналған диетатөменде сипатталған басқа тамақтану бойынша кеңестер арасында адамдар ұзақ және сау өмір сүре алады. Дәстүрлі диеталар жиі қиын және стресстік болуы мүмкін, бірақ ұзақ өмір диета жоспары көптеген адамдарға қолайлы болуы мүмкін практикалық және бірегей диеталық бағдарлама.
Доктор Алекс Хименес DC, CCST Insight
Ұзақ өмір сүру диета жоспарының 8 тамақтану туралы кеңестері
Пескара диетасын ұстаныңыз
Ұзақ өмірге арналған диета жоспарының бөлігі ретінде пескара диетасын ұстаныңыз, ол шамамен 100 пайыз өсімдіктер мен балықтарға негізделген. Сондай-ақ, балықты тұтынуды апта сайын екі-үш порциямен шектеп, тунца, қылыш, скумбрия және галибут тәрізді құрамында сынаптың жоғарылауы бар балықтардан аулақ болыңыз. Егер сіз 65-ден асып жатсаңыз және аздаған бұлшық массасын, күш пен майды бастан бастасаңыз, фита немесе печорино сияқты жұмыртқаны және арнайы ірімшіктерді қоса, жануарларға негізделген басқа да тағамдарды қоса, диетаңызға көп балық қосыңыз, және ешкіден жасалған йогурт сүт.
Тым көп ақуызды жеуге болмайды
Ұзақ мерзімді диета жоспары бойынша біз күн сайын дене салмағының фунтына 0.31-ден 0.36 грамм ақуызды жеуіміз керек. Егер сіз 130lб салмағыңыз келсе, 40-ге шамамен 47 грамм ақуызды жеуге тура келеді күні, немесе 1.5 лосось, 1 кесе ноталар немесе 2 1 / 2 кәмпиттерінің баламасы, оның ішінде 30 грамм бір тағаммен тұтынылуы тиіс. Егер сіз 200-ді 220lб-қа салмақ болсаңыз, күніне 60-ден 70 грамм ақуызға немесе лосось, 3 1 / 2 шыныаяқтың немесе 1 1 / 2 нопалардың шыны аяқтарына эквивалентті. Протеинді тұтыну 65 жасынан кейін артуы керек. Көпшілігіміз үшін 10 дейін 20 пайызға дейін немесе 5-ден күн сайын 10 граммға дейін жетеді. Ақырында, ұзақ өмір сүру диета балықтың жануарлар ақуыздарын қоспағанда, қызыл ет, ақ ет және құс сияқты жануарлар белоктарынан бос. Бұл бірегей диеталық бағдарлама денсаулық пен сауықтылықты оңтайландыру үшін бұршақ пен жаңғақ сияқты өсімдік белоктарында салыстырмалы түрде жоғары.
Жақсы майлар мен кешенді көмірсуларды көбейту
Ұзақ мерзімді диета жоспарының бөлігі ретінде лосось, бадам, жаңғақ және зәйтүн майы сияқты көп мөлшерде полиқанықпаған майларды көп мөлшерде жеуге тура келеді, ал сіз аз мөлшерде қаныққан, гидрирленген және транзит майларын жеуге тиіссіз. Сонымен қатар, ұзақ өмір сүру диета жоспарының бөлігі ретінде сіз тұтас бидай наны, бұршақ және көкөністерде кездесетін күрделі көмірсуларды жеуіңіз керек. Пастаны, күрішті, нан, жеміс-жидекті және жеміс шырындарын шектеуді ұмытпаңыз, олар ішекке жеткен кезде қантқа айналуы мүмкін.
Диеталық қосымшаларды алыңыз
Адам ағзасына ақуыз, омега-ХНУМХ және омега-ХНУМХ сияқты маңызды май қышқылдары, витаминдер, минералдар, тіпті дұрыс жұмыс істеу үшін қант қажет. Белгілі бір қоректік заттардың тұтынылуы тым төмен болған кезде, адам денесінің жөндеу, ауыстыру және қорғаныс әдістері саңырауқұлақтар, бактериялар мен вирустарға зиян келтіруі мүмкін денсаулықты әртүрлі мәселелерге әкеп соғуы мүмкін. Сіздің денсаулық сақтау маманыңыз ұсынғандай, әсіресе omega-3 үшін витамин мен минералды тағамдық қоспаларды алыңыз.
Әртүрлі азық-түліктерді сіздің А-дан алыңызата-анасы
Сізге керекті қоректік заттардың барлығын алу үшін, сізге көптеген тағамдарды жеуге тура келеді, бірақ ата-аналарыңыз, әжелеріңіз бен әжелеріңіз бен ата-әжелеріңіздің әдеттегі тағамдарын таңдауыңыз керек. Мысалға, сүт тұтынатын көптеген солтүстік Еуропа елдерінде лактозаның төзімсіздігі салыстырмалы түрде сирек кездеседі, алайда лактозаның төзімсіздігі ертедегі дәстүрлі диетаның тарихи бөлігі болмаған оңтүстік еуропалық және азиялық елдерде кеңінен таралған. Егер Америка Құрама Штаттарында тұратын жапон ертегілерінің біреуі күтпеген жерден әжесі мен әжелерінің асханасында сирек кездесетін сүтті ішуді шешсе, олар ауырып қалады. Мұндай жағдайларда жиі кездесетін проблемалар төзімсіздік немесе аутоэммуния, мысалы, целий ауруы бар адамдарда кездесетін нан мен макарон сияқты глютенге бай тағамға жауап ретінде. Дәлелдемелер қажет болса да, азық-түлік төзімсіздіктері көптеген аутоиммундық бұзылулар, соның ішінде қант диабеті, колит және Crohn ауруымен байланысты болуы мүмкін.
Күніне екі тағам мен тағамдарды жегіңіз
Ұзақ өмірлік диета жоспарына сәйкес күн сайын таңғы асты және бір үлкен тағамды, сонымен қатар аз калориялы, қант деңгейі төмен тағамдық тағамдарды ішу өте қолайлы. Кейбір адамдар үшін күн сайын үш рет тамақ ішіп, тамақ ішуге кеңес берілуі мүмкін. Көптеген тамақтану нұсқаулықтары күн сайын бес-алты тамақтануға кеңес береді. Адамдарға жиі тамақтануға кеңес бергенде, оларға калориялардың тұтынылуын реттеу қиынға соғуы мүмкін. Соңғы жиырма жыл ішінде Америка Құрама Штаттарындағы халықтың шамамен 70 пайызы артық салмақ немесе семіздік деп саналады. Ұзақ өмірлік диета жоспарын шамадан тыс жеу әлдеқайда қиын, егер сіз күн сайын тек екі жарым тамақ ішсеңіз. Салмақ қосуға әкелетін мөлшерге жету үшін бұршақ тұқымдастардың, көкөністер мен балықтардың жаппай бөліктерін қажет етеді. Тамақтанудың жоғары мөлшері, сонымен қатар тамақ мөлшері сіздің асқазаныңыз бен миыңызға сізде жеткілікті тамақ болғандығы туралы сигнал береді. Ас қорыту мәселелерін болдырмау үшін осы бір негізгі тамақтану жүйесін кейде екі тағамға бөлуге тура келеді. Арықтауға бейім ересектер мен қарт адамдар күніне үш рет тамақтануы керек. Артық салмақтан арылуға тырысатын адамдар үшін, сондай-ақ артық салмақ немесе семіздік үшін тамақтанудың ең жақсы кеңесі күнделікті таңғы асты ішу керек; кешкі немесе түскі ас ішіңіз, бірақ екеуін де қабылдамаңыз, және 100 калориядан аз және 3 - 5 г қанттан аспайтын бір тағаммен ауыстырыңыз. Қай тамақтан бас тарту сіздің өмір салтыңызға байланысты, дегенмен денсаулығына байланысты таңғы астан бас тарту ұсынылмайды. Түскі астан бас тартудың артықшылығы - бос уақыт пен күш. Бірақ үлкен кешкі ас ішудің, әсіресе қышқыл рефлюксінен немесе ұйқы проблемасынан зардап шегетін адамдар үшін жетіспеушілік бар. Алайда кешкі астан бас тартудың жетіспеушілігі - бұл олардың күнделікті әлеуметтік тамағын жоятындығында.
Күн сайын 12 сағаттық терезеде жегіңіз
Көптеген жүздеген жастар қабылдаған тағы бір әдеттегі тамақ әдеті күн сайын 12 сағаттық терезедегі барлық тамақ пен тағамдармен шектеледі немесе шектейді. Бұл әдіс тиімділігі адам және жануарлардың зерттеу жұмыстарында да дәлелденді. Әдетте, сіз 8-да таңғы асты жеп, кешкі асты 8-да тамақтандырыңыз. Он сағаттық немесе одан да аз уақытты тамақтанатын терезе салмақ жоғалту үшін одан да жақсы болуы мүмкін, бірақ оны сақтау әлдеқайда қиын, ал өт тастар сияқты жанама әсерлердің пайда болу қаупін арттырады және жүрек-қан тамырлары ауруларының даму мүмкіндігін арттырады. Ұйықтаудан үш-төрт сағат бұрын жеуге болмайды.
ProLon диета ұстаныңыз
ХNUMX жасқа толған салауатты адамдар оны ұстануы керек ProLon ораза ұстаушыларға арналған диета, 5 күндік тамақтану бағдарламасы кем дегенде жылына екі рет. FMD ұзақ өмір сүру диета жоспары ұсынған негізгі принциптердің бірі болып табылады. Аштықты емдейтін диета оразаның орасан зор пайдасы үшін шынымен ораза ұстамайды. 800-ден нақты мөлшерде және әрбір күн үшін таңбаланған азық-түліктердің комбинациясы кезінде 1,100 калориясын тамақтандыру арқылы сіз адам денесін «ораза» жағдайына «алдамай» аласыз. Доктор Вальтер Лонго әртүрлі зерттеулердің арқасында тағамның денесін осы жолмен айыра отырып, біздің клеткаларымыз ішкі тіндерді бұзып, қалпына келтіру, өлтіру және ауыстыру, қалпына келтіру, зақымдалған клеткаларды жою арқылы бастайды. Сонымен қатар, ораза әртүрлі денсаулық мәселелеріне кері әсерін тигізуі мүмкін, рак клеткаларын жоюға және Альцгеймер ауруы даму мүмкіндігін елеулі түрде төмендетуге мүмкіндік береді.
Доктор Вальтер Лонгоның кітабында ұсынылған ұзақ өмір диеталық жоспарымен сіз жақсы тамақтанасыз, жақсы сезінесіз және бұл салмақ жоғалту жоспары болмаса да, сіз бірнеше фунт тастай аласыз. Сізге осы ерекше диеталық бағдарламаның көмегімен күрделі тамақтану ережелерін қарастырудың қажеті жоқ және қиын таңдау жасау керек. Осы өмір салтын өзгерткеннен кейін сіз өзіңіздің денсаулығыңыз бен денсаулығыңызды жақсартасыз сіздің ұзақ өмір сүру. The Біздің ақпаратымыздың көлемі широпротикамен, жұлын денсаулығымен және функционалды медицинамен шектеледі. Тақырыпты әрі қарай талқылау үшін д-р Алекс Хименеске немесе бізбен хабарласыңыз 915-850-0900 .
Доктор Алекс Хименес мырзаның жетекшілігімен
Қосымша тақырыпты талқылау: Жедел арқа ауруы
Арқа ауруы - мүгедектіктің ең көп таралған себептерінің бірі және бүкіл әлем бойынша жұмыстан босатылған күндер. Кері ауруы дәрігерлік кеңселерге баратын ең көп тараған себептерге байланысты, тек жоғарғы тыныс жолдарының инфекцияларымен шектеледі. Шамамен ХNUMX пайызы халық өмірінде кем дегенде бір рет ауырсынуды сезінеді. Сіздің омыртқаның - бұл жұмсақ маталардан тұратын сүйек, буын, байлам және бұлшықеттерден тұратын күрделі құрылым. Жарақат және / немесе ауырлататын жағдайлар сияқты қатты дискілер, ақыр соңында арқа ауырсыну белгілеріне алып келуі мүмкін. Спорттық жарақаттар немесе автомобиль апатының жарақаттары көбінесе арқадағы ауырсынудың жиі себебі болып табылады, бірақ кейде ең қарапайым қозғалыстың ауыр нәтижесі болуы мүмкін. Бақытымызға орай, широпрактикалық күтім сияқты альтернативті емдеу параметрлері омыртқаның түзетуін және манипуляцияларды қолдану арқылы ауырсынуды жеңілдетуге көмектеседі, ақыр соңында ауруды жеңілдетеді.
XYMOGEN s Эксклюзивті кәсіби формулалар таңдаулы лицензиялық денсаулық сақтау мамандары арқылы қол жетімді. Интернетті сату және XYMOGEN формулаларын дисконттау қатаң тыйым салынады.
Мақтанышпен, Александр Хименес XYMOGEN формулаларын біздің қамқорлығымыздағы науқастарға ғана қол жетімді етеді.
Шұғыл қол жеткізу үшін дәрігерге кеңес беру үшін біздің кеңсемізге хабарласыңыз.
Егер сіз науқас болсаңыз Медициналық және хиропрактикалық жарақаттар клиникасы, сіз қоңырау шалу арқылы XYMOGEN туралы біле аласыз 915-850-0900.
Сізге ыңғайлы болу үшін және шолу XYMOGEN өнімдері төмендегі сілтемемен танысыңыз. *XYMOGEN-Каталог -жүктеу
* Жоғарыда аталған барлық XYMOGEN саясаты қатаң сақталады.
IFM's Find A Practitioner құралы пациенттерге әлемнің кез келген жерінде функционалдық медицина мамандарын табуға көмектесу үшін жасалған функционалдық медицинадағы ең үлкен бағыттаушы желі болып табылады. IFM сертификатталған тәжірибешілері олардың функционалдық медицина бойынша кең білімін ескере отырып, іздеу нәтижелерінде бірінші болып табылады.