Артқа клиниканың тотығу стрессі хиропротикалық және функционалдық медицина тобы. Тотығу стрессі реактивті оттегі (бос радикалдар) өндірісі мен антиоксиданттық қорғаныс арасындағы тепе-теңдіктің бұзылуы ретінде анықталады. Басқаша айтқанда, бұл бос радикалдардың өндірісі мен антиоксиданттар арқылы бейтараптандыру арқылы ағзаның зиянды әсерге қарсы тұру немесе детоксикациялау қабілеті арасындағы теңгерімсіздік. Тотығу стрессі ағзадағы көптеген патофизиологиялық жағдайларға әкеледі. Оларға нейродегенеративті аурулар, яғни Паркинсон ауруы, Альцгеймер ауруы, гендік мутациялар, қатерлі ісіктер, созылмалы шаршау синдромы, нәзік X синдромы, жүрек және қан тамырларының бұзылуы, атеросклероз, жүрек жеткіліксіздігі, жүрек соғысы және қабыну аурулары жатады. Тотығу бірнеше жағдайларда жүреді:
жасушалар қуат алу үшін глюкозаны пайдаланады
иммундық жүйе бактериялардан сақтанып, қабынуды тудырады
органдардың ластаушы заттарды, пестицидтерді және темекі түтіндерін нашарлатады
Біздің органдарымызда кез-келген уақытта тотығуға алып келетін миллиондаған процестер бар. Төменде бірнеше белгілер бар:
Шаршау
Жадтың жоғалуы немесе мидың тұмауы
Бұлшықет немесе бірлескен ауырсыну
Сұр шаштармен қатар әжімдер
Көру қабілетінің төмендеуі
Бас ауруы және шуылдың сезімталдығы
Инфекцияға сезімталдық
Органикалық тағамдарды таңдау және қоршаған ортаңыздағы токсиндерден аулақ болу үлкен айырмашылықты тудырады. Бұл стрессті азайтумен қатар тотығуды азайтуға пайдалы болуы мүмкін.
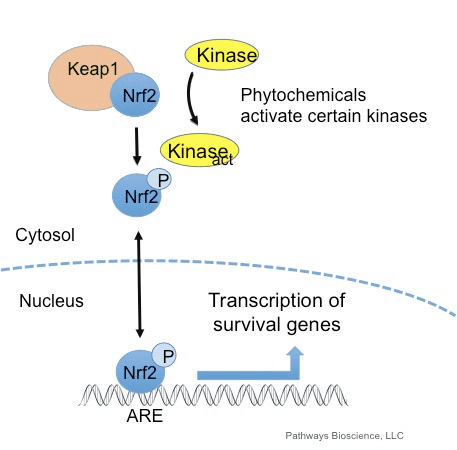
Тотықтырғыштар әдетте адам ағзасындағы маңызды процестерді, соның ішінде жасушалардың бөлінуін, қабынуды, иммундық функцияны, аутофагияны және стресске жауап беруді реттеу үшін бақыланатын түрде шығарылады. Дегенмен, осы тотықтырғыштардың бақылаусыз өндірісі ықпал етуі мүмкін тотықтырғыш стресс, бұл уыттылықтың, созылмалы аурулардың және қатерлі ісіктердің дамуына әкелетін жасушалық функцияға әсер етуі мүмкін. Адам ағзасының антиоксиданттық қорғаныс механизмдері жасушаның тотықтырғыштарға реакциясын бақылайтын өмірлік маңызды жолдар тізбегі арқылы реттеледі. Ядролық фактор эритроид 2-байланысты фактор, басқаша Nrf2 деп аталады, тотықтырғыштарға жасушалық төзімділіктің дамып келе жатқан реттегіші болып табылады. Төмендегі мақаланың мақсаты - митохондриялық функциядағы Nrf2 рөлін талқылау және көрсету.
дерексіз
Транскрипция факторы NF-E2 p45 байланысты фактор 2 (Nrf2; ген атауы NFE2L2) антиоксидантты, қабынуға қарсы және детоксикация ферменттерін қоса, цитопротекторлық ақуыздардың әртүрлі желілерінің гендік экспрессиясын реттеу арқылы стресс жағдайында бейімделуге және өмір сүруге мүмкіндік береді. зақымдалған макромолекулаларды қалпына келтіруге немесе жоюға көмектесетін ақуыздар ретінде. Nrf2 глутатион, тиоредоксин және NADPH биосинтезін, пайдалануын және регенерациясын реттеу және митохондрия мен NADPH оксидаза арқылы реактивті оттегі түрлерінің өндірісін бақылау арқылы жасушалық тотығу-тотықсыздану гомеостазын қолдауда шешуші рөл атқарады. Гомеостатикалық жағдайларда Nrf2 митохондриялық мембрана потенциалына, май қышқылының тотығуына, тыныс алу үшін субстраттардың (NADH және FADH2/сукцинат) қолжетімділігіне және ATP синтезіне әсер етеді. Стресс жағдайында немесе өсу факторын ынталандыру жағдайында Nrf2 активтенуі митохондриядағы реактивті оттегі түрлерінің өндірісінің жоғарылауына 3-ші ақуызды транскрипциялық реттеу арқылы қарсы әсер етеді және 1 ядролық тыныс алу факторы мен пероксисома пролифераторы арқылы белсендірілген рецептор деңгейін ұстап тұру арқылы митохондрия биогенезіне әсер етеді? коактиватор 1?, сондай-ақ пурин нуклеотидтерінің биосинтезін ынталандыру арқылы. Табиғи изотиоцианат сульфорафан сияқты фармакологиялық Nrf2 белсендіргіштері тотықтырғыш арқылы митохондрия өткізгіштігінің өту тесігінің ашылуын және митохондрияның ісінуін тежейді. Бір қызығы, бастапқыда Nrf1,4 активаторы ретінде жасалған синтетикалық 1,2,3-дифенил-2-триазол қосылысы митофагияны дамытады, осылайша жалпы митохондриялық гомеостазға ықпал етеді. Осылайша, Nrf2 митохондриялардың құрылымдық және функционалдық тұтастығын қолдаудағы көрнекті ойыншы болып табылады және бұл рөл стресс жағдайында өте маңызды.
Түйінді сөздер:Биоэнергетика, Цитопротекция, Keap1, Митохондрия, Nrf2, Бос радикалдар
Highlights
Nrf2 жасушалық тотығу-тотықсыздану гомеостазын сақтауда шешуші рөл атқарады.
Nrf2 митохондриялық мембрана потенциалына және ATP синтезіне әсер етеді.
Nrf2 митохондриялық май қышқылының тотығуына әсер етеді.
Nrf2 митохондриялардың құрылымдық және функционалдық тұтастығын қолдайды.
Nrf2 активаторлары митохондриялық функция бұзылған кезде пайдалы әсер етеді.
кіріспе
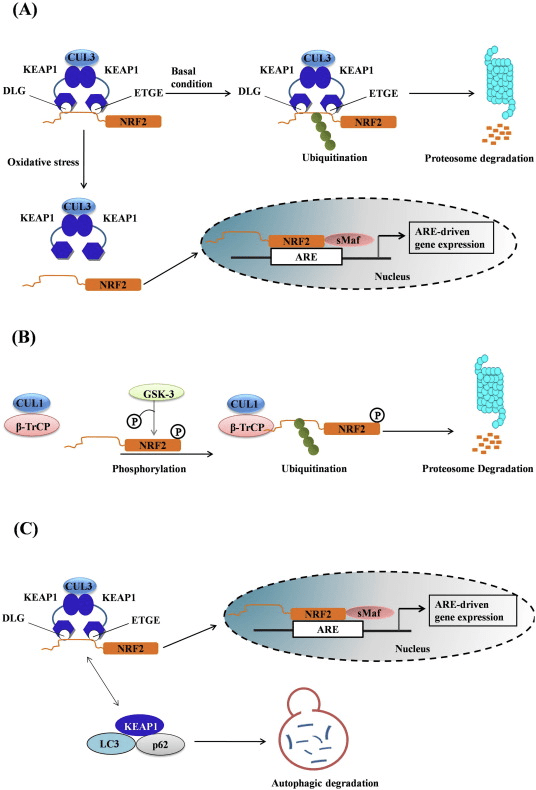
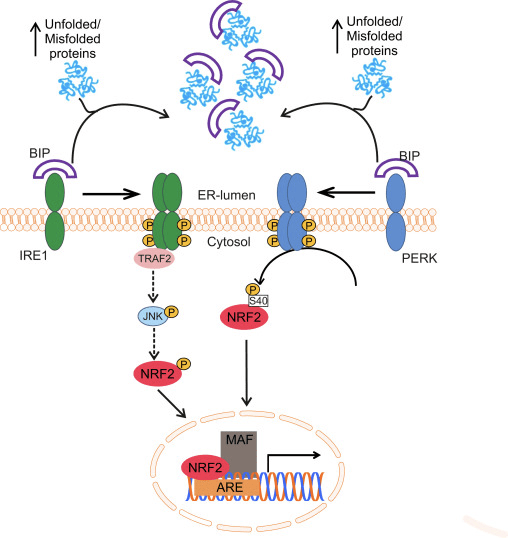
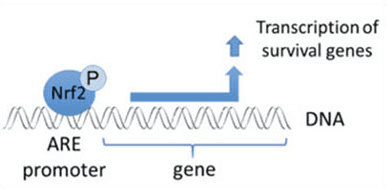
Транскрипция факторы NF-E2 p45 байланысты фактор 2 (Nrf2; ген атауы NFE2L2) әртүрлі цитопротекторлық белсенділіктері бар белоктарды кодтайтын гендер желілерінің экспрессиясын реттейді. Nrf2 өзі ең алдымен ақуыз тұрақтылығы деңгейінде бақыланады. Базальды жағдайларда Nrf2 үздіксіз убиквитинацияға және протеазомалық ыдырауға ұшырайтын қысқа мерзімді ақуыз болып табылады. Nrf2 деградациясына ықпал ететін үш белгілі ubiquitin ligase жүйесі бар. Тарихи түрде Nrf2 бірінші теріс реттеушісі табылған Келх тәрізді ECH-ассоциацияланған протеин 1 (Keap1) [1], Куллин 3 (Cul3)/Rbx1 убиквитин лигазасының субстрат адаптер ақуызы [2], [3], [ 4]. Keap1 ubiquitination және протеазомалық деградация үшін Nrf2-ге бағытталған жоғары тиімді циклдік механизмді пайдаланады, оның барысында Keap1 циклдің жалғасуына мүмкіндік беретін үздіксіз регенерацияланады (1А-сурет) [5]. Nrf2 сонымен қатар гликоген синтаза киназасы (GSK)3/?-TrCP-тәуелді Cul1 негізіндегі убиквитин лигазасы арқылы деградацияға ұшырайды [6], [7]. Жақында эндоплазмалық ретикулум кернеуі жағдайында Nrf2 E3 ubiquitin лигаза Hrd1 делдалдық процесте убиквитинацияланады және ыдырайтыны туралы хабарланды [8].
Сурет 1 Nrf1-нің Keap2-делдалдық деградациясы үшін циклдік дәйекті байланысу және регенерация үлгісі. (A) Nrf2 бос Keap1 димерімен дәйекті түрде байланысады: алдымен жоғары аффинді ETGE (қызыл таяқшалар) байланыстыру домені арқылы, содан кейін төмен аффинді DLG (қара таяқшалар) байланыстырушы домен арқылы. Ақуыз кешенінің бұл конформациясында Nrf2 убиквитинациядан өтеді және протеазомалық деградацияға бағытталған. Бос Keap1 қалпына келтіріліп, жаңадан аударылған Nrf2-ге қосыла алады және цикл қайтадан басталады.(B) Индукторлар (ақ гауһар) Keap1 сенсорлық цистеиндерімен (көк таяқшалар) әрекеттеседі, бұл конформациялық өзгеріске және субстрат адаптерінің белсенділігінің бұзылуына әкеледі. Бос Keap1 регенерацияланбайды, ал жаңадан синтезделген Nrf2 жинақталып, ядроға ауысады.
Ubiquitin-ligase субстрат адаптері ақуызы ретінде қызмет етуден басқа, Keap1 Nrf2 шағын молекулалы активаторларының кең ауқымы үшін сенсор болып табылады (индукторлар деп аталады) [9]. Индукторлар Keap1 [2], [1] ішіндегі спецификалық цистеин қалдықтарын химиялық түрлендіру арқылы немесе Keap10:Nrf11 байланыстыру интерфейсін [1], [2] тікелей бұзу арқылы Nrf12-нің Keap13 арқылы ыдырау циклін блоктайды. Демек, Nrf2 ыдырамайды, ал транскрипция факторы жинақталып, ядроға ауысады (1В-сурет), онда шағын Maf протеині бар гетеродимер түзеді; антиоксиданттық-жауап элементтерімен, оның мақсатты гендерінің жоғарыдағы реттеуші аймақтарымен байланысады; және транскрипцияны бастайды [14], [15], [16]. Nrf2 нысаналарының батареясы ксенобиотикалық метаболизмнің ферменттерін, антиоксидантты және қабынуға қарсы функциялары бар ақуыздарды және протеазомалық суббірліктерді, сондай-ақ жасушалық тотығу-тотықсыздану гомеостазын реттейтін және аралық метаболизмге қатысатын ақуыздарды қоса алғанда, әртүрлі цитопротекторлық функциялары бар ақуыздардан тұрады.
Nrf2: жасушалық тотығу-тотықсыздану гомеостазының негізгі реттеушісі
Nrf2 функциясы жасушалық тотығу-тотықсыздану гомеостазының негізгі реттеушісі ретінде кеңінен танылған. ?-глутамилцистеин лигазасының каталитикалық және реттеуші бөлімшелерінің гендік экспрессиясы, төмендетілген глутатион (GSH) биосинтезіндегі жылдамдықты шектейтін сатыны катализдейтін фермент, Nrf2 [17] арқылы тікелей реттеледі. Цистинді жасушаларға импорттайтын xc- жүйесінің xCT суббірлігі де Nrf2 [18] тікелей транскрипциялық нысанасы болып табылады. Жасушада цистин GSH биосинтезінің прекурсоры болып табылатын цистеинге айналады. GSH биосинтезіндегі рөлінен басқа, Nrf2 глутатион редуктаза 1 [19], [20] координацияланған транскрипциялық реттеу арқылы глутатионды төмендетілген күйде ұстауға арналған құралдарды қамтамасыз етеді, ол NADPH-тен қалпына келтіретін эквиваленттерді пайдалана отырып, тотыққан глутатионды GSH-ға дейін төмендетеді. . Қажетті NADPH төрт негізгі NADPH-генерациялайтын ферменттермен қамтамасыз етіледі, алма ферменті 1 (ME1), изоцитратдегидрогеназа 1 (IDH1), глюкоза-6-фосфатдегидрогеназа (G6PD) және 6-фосфоглюконатдегидрогеназа (PGD), олардың барлығы ішінара Nrf2 (2-сурет) [21], [22], [23], [24] арқылы транскрипциялық реттеледі. Бір қызығы, Nrf2 сонымен қатар NAD(P)H түзетін кофактор ретінде NAD(P)+ қолданатын альдегиддегидрогеназаның [25] цитозолдық, микросомалық және митохондриялық формаларының индукцияланатын ген экспрессиясын реттейді. Шынында да, NADPH деңгейлері және NADPH/NADP+ қатынасы Nrf2-нокаут (Nrf2-KO) тышқандарынан оқшауланған эмбриондық фибробласттарда олардың жабайы типтегі (WT) аналогтары жасушаларымен салыстырғанда төмен, ал NADPH деңгейлері Nrf2 нокаутында төмендейді. конститутивті белсенді Nrf2 бар қатерлі ісік жасушаларының сызықтары [26]. Күтілгендей, GSH деңгейі Nrf2 бұзылған жасушаларда төмен; керісінше, генетикалық немесе фармакологиялық жолмен Nrf2 белсендіру GSH жоғарылауына әкеледі [27], [28], [29]. Маңыздысы, Nrf2 сонымен қатар маңызды болып табылатын тиоредоксин [30], [31], [32], тиоредоксин редуктаза 1 [28], [29], [32], [33] және сульфиредоксин [34] генінің экспрессиясын реттейді. тотыққан ақуыз тиолдарын қалпына келтіру үшін.
2-сурет Жылдам көбейетін жасушалардың метаболизміндегі Nrf2 рөлі. Nrf2 тотығу иініндегі [яғни, глюкоза-6-фосфатдегидрогеназа (G6PD) және 6-фосфоглюконатдегидрогеназа (PGD)] және тотықтырмайтын тармақтағы [яғни, трансальдолаза 1 (TALDO1) және транскетолаза (яғни, қышқылдану иініндегі) ферменттерді кодтайтын гендердің оң реттеушісі болып табылады. TKT)] пентозофосфат жолының. G6PD және PGD NADPH жасайды. Nrf2 сонымен қатар NADPH-генерациялайтын басқа екі ферменттің, алма ферментінің 1 (ME1) және изоцитратдегидрогеназа 1 (IDH1) генінің экспрессиясын реттейді. Де жаңа пуриндік биосинтетикалық жолға кіруді катализдейтін фосфорибозилпирофосфат амидотрансферазасының (PPAT) гендік экспрессиясы да Nrf2 арқылы оң реттеледі, сондай-ақ метилентетрагидрофолат дегидрогеназа 2 (MTHFD2), амитохрондағы критикалық рөл атқарады. де жаңа пурин биосинтезі үшін бір көміртекті бірліктерді қамтамасыз ету. Пируваткиназа (PK) Nrf2 арқылы теріс реттеледі және гликолитикалық аралық өнімдердің түзілуіне және G6PD-мен бірге пентозофосфат жолы арқылы метаболиттердің өтуіне және нуклеин қышқылдарының, аминқышқылдарының және фосфолипидтердің синтезіне ықпал етеді деп күтілуде. Nrf2 ATP-цитрат-лиаза (CL) генінің экспрессиясын теріс реттейді, бұл цитраттың митохондриялық утилизация үшін немесе (изоцитрат арқылы) IDH1 үшін қолжетімділігін арттыруы мүмкін. Қызыл және көк сәйкесінше оң және теріс реттеуді көрсетеді. Митохондрия сұр түспен көрсетілген. Метаболиттің аббревиатуралары: G-6-P, глюкоза 6-фосфат; F-6-P, фруктоза 6-фосфат; F-1,6-BP, фруктоза 1,6-бисфосфат; ГА-3-Р, глицеральдегид 3-фосфат; 3-ПГ, 3-фосфоглицерат; ПЭП, фосфоэнолпируват; 6-P-Gl, 6-фосфоглюконолактон; 6-ПГ, 6-фосфоглюконат; R-5-P, рибулоза 5-фосфат; ПРПП, 5-фосфорибозил-?-1-пирофосфат; THF, тетрагидрофолат; IMP, инозинмонофосфаты; АМФ, аденозинмонофосфаты; GMP, гуанозин монофосфаты.
Nrf2-нің жасушалық тотығу-тотықсыздану гомеостазының басты реттеушісі ретіндегі шешуші рөлін ескере отырып, WT жасушаларымен салыстырғанда, Nrf2 бұзылған жасушаларда реактивті оттегі түрлерінің (ROS) жоғары болуы таңқаларлық емес (Nrf2-KO) [35]. Бұл айырмашылық әсіресе тотығу стресін тудыратын агенттермен күресу кезінде таң қалдырады. Сонымен қатар, Nrf2 жетіспейтін жасушалар әртүрлі түрдегі тотықтырғыштардың уыттылығына анағұрлым сезімтал және бірдей жағдайларда WT жасушаларын тиімді және ұзақ мерзімді қорғауды қамтамасыз ететін Nrf2 индукторларымен қорғала алмайды [29], [36] , [37]. Жалпы жасушалық тотығу-тотықсыздану гомеостазынан басқа, Nrf2 митохондриялық тотығу-тотықсыздану гомеостазын қолдау үшін де маңызды. Осылайша, WT-мен салыстырғанда, жалпы митохондриялық NADH пулы Keap1-KO-да айтарлықтай артады және Nrf2-KO жасушаларында күрт төмендейді [35].
Тірі жасушаларды бейнелеуді қолдана отырып, біз жақында WT, Nrf2-KO немесе Keap1-knockdown (Keap1-KD) тышқандарынан бөлінген бастапқы глионерональды кокультуралар мен ми тінінің кесінділеріндегі ROS өндірісінің жылдамдығын бақыладық [38]. Күтілгендей, ROS өндірісінің жылдамдығы олардың WT аналогтарымен салыстырғанда Nrf2-KO жасушалары мен тіндерінде жылдамырақ болды. Дегенмен, біз WT және Keap1-KD генотиптері арасындағы айырмашылықтың шамасы WT және Nrf1-KO арасындағы айырмашылыққа қарағанда аз болғанымен, WT-мен салыстырғанда, Keap2-KD жасушаларында ROS өндірісінің жоғары қарқыны бар екенін күтпеген байқау жасадық. . Содан кейін біз NOX2 және NOX4 мРНҚ деңгейлерін, ми патологиясына қатысы бар екі NADPH оксидаза (NOX) изоформасының каталитикалық бөлімшелерін талдадық және Nrf2 тапшылығы жағдайында NOX2 күрт артқанын, ал Nrf4 болғанда NOX2 жоғарылайтынын анықтадық. аз дәрежеде болса да конститутивтік белсендіріледі. Сандық тұрғыдан алғанда, мутантты тышқандардың жасушалары мен тіндеріндегі жоғарылаудың шамасы ROS өндірісінің сәйкес өсуіне параллельді [38]. Бір қызығы, Nrf2 NADPH оксидазасын реттеп қана қоймайды, бірақ NADPH оксидазасы шығаратын ROS өкпе эпителий жасушаларында және кардиомиоциттерде көрсетілгендей Nrf2 белсендіреді [39], [40]. Сонымен қатар, жақында жүргізілген зерттеу Nrf2-нің NADPH оксидазасына тәуелді активтенуі митохондриялық зақымданудан және созылмалы қысымның шамадан тыс жүктелуі кезінде жүректегі жасушалардың өлімінен қорғаудың маңызды эндогендік механизмін құрайтынын көрсетті [41].
NADPH оксидазасының каталитикалық белсенділігінен басқа, митохондриялық тыныс алу ROS-тың тағы бір негізгі жасушаішілік көзі болып табылады. Митохондрияға спецификалық MitoSOX зондын қолдану арқылы біз бастапқы глионероналды кокультуралардағы жалпы ROS өндірісіне митохондриялық текті ROS үлесін зерттедік. WT, Nrf2-KO немесе Keap1-KD тышқандарынан [38]. Күтілгендей, Nrf2-KO жасушаларында митохондриялық ROS өндірісінің WT қарағанда жоғары қарқыны болды. Жалпы ROS өндірісінің нәтижелерімен келісе отырып, Keap1-KD митохондриялық ROS өндірісінің жылдамдығы WT жасушаларымен салыстырғанда жоғары болды. Маңыздысы, I кешенін ротенонмен блоктау WT және Keap1-KD жасушаларында митохондриялық ROS өндірісінің күрт артуына әкелді, бірақ Nrf2-KO жасушаларына әсер етпеді. Пируват қосылғаннан кейін WT жасушаларында митохондриялық ROS өндірісінің күтілетін ұлғаюынан айырмашылығы (NADH қолжетімділігін арттыру, митохондриялық мембрана потенциалын арттыру және тыныс алуды қалыпқа келтіру үшін) Nrf2-KO жасушаларында ROS өндірісі төмендеді. Бұл тұжырымдар бірге Nrf2 болмаған жағдайда: (i) I комплексінің белсенділігі бұзылғанын, (ii) I комплексінің белсенділігінің бұзылуы субстраттардың шектелуіне байланысты және (iii) кешеннің бұзылған белсенділігін көрсетеді. I - митохондриялық ROS өндірісінің жоғарылауының негізгі себептерінің бірі, мүмкін II комплексінен кері электрон ағынының арқасында.
Nrf2 митохондриялық мембрананың потенциалы мен тыныс алуына әсер етеді
Митохондриялық мембрана потенциалы (??м) митохондрияның денсаулығы мен жасушаның зат алмасу күйінің әмбебап көрсеткіші болып табылады. Сау жасушада ??m митохондриялық тыныс алу тізбегі арқылы сақталады. Бір қызығы, эстроген рецепторлары теріс адам сүт безі эпителийінің MCF10A жасушалық сызығында культура негізіндегі протеомика зерттеуіндегі аминқышқылдарымен тұрақты изотоптық таңбалау митохондриялық электрондарды тасымалдау тізбегі NDUFA4 компонентінің фармакологиялық белсендірумен (NDUFA2, сульфоррафан2 арқылы) жоғары реттелетінін көрсетті. ал Nrf1-нің генетикалық жоғарылауы (Keap2 нокдаун арқылы) цитохром с оксидазасының COX4 және COX1I42 суббірліктерінің төмендеуіне әкеледі [2]. Екі өлшемді гельдік электрофорез және матрицалық лазерлік десорбция/ионизациялық масс-спектрометрия көмегімен бауыр протеомын зерттеу Nrf43 ATP синтаза суббірлігінің экспрессиясын реттейтінін анықтады. [1]. Сонымен қатар, I кешенінің [44] белсенділігін қолдауда рөл атқаратын митохондриялық ақуыз DJ-2, фармакологиялық немесе генетикалық белсендірудің нейропротекторлық әсерлеріне қарамастан, Nrf45 [46], [2] тұрақтандыруы туралы хабарланған. Nrf1 саны DJ-47-ге тәуелсіз [XNUMX]. Дегенмен, бұл бақылаулардың митохондриялық функцияға әсері зерттелмеген.
Nrf2 тапшылығы жағдайында I комплексінің белсенділігінің бұзылуына сәйкес, Nrf2-KO тышқанның эмбриондық фибробласттарында (MEFs) және өсірілген бастапқы глионейрондық жасушаларда олардың WT аналогтарымен салыстырғанда базальді ??m төмен болады (3-сурет, кірістірілген). [35]. Керісінше, Nrf2 генетикалық конститутивтік жоғары реттелгенде (Keap1-ді нокдаун немесе нокаут арқылы) базальды ??m жоғарырақ болады. Генотиптер арасындағы бұл ??m айырмашылықтары тыныс алудың Nrf2 белсенділігінің әсер ететінін көрсетеді. Шынында да, базальды күйдегі оттегін тұтынуды бағалау WT-мен салыстырғанда, оттегі тұтынуы Nrf2-KO және Keap1-KO MEF-де тиісінше ~50 және ~35% төмен екенін көрсетті.
Сурет 3 Nrf2 тапшылығы жағдайында бұзылған митохондриялық функцияның ұсынылатын механизмі. (1) ME1, IDH1, G6PD және PGD деңгейлерінің төмендеуі NADPH деңгейлерінің төмендеуіне әкеледі. (2) GSH деңгейі де төмен. (3) ME1 белсенділігінің төмендігі митохондрияға түсетін пируват пулын азайтуы мүмкін. (4) NADH генерациясы баяу, бұл I кешенінің белсенділігінің бұзылуына және митохондриялық ROS өндірісінің жоғарылауына әкеледі. (5) Митохондриялық белоктардағы FAD-ның FADH2-ге дейін төмендеуі де азаяды, FADH2-ден UbQ-ға және III комплекске электрон ағынын төмендетеді. (6) UbQH2 баяу түзілуі сукцинатдегидрогеназа ферментінің белсенділігін төмендетуі мүмкін. (7) ROS деңгейінің жоғарылауы II кешеннің белсенділігін одан әрі тежеуі мүмкін. (8) Май қышқылдарының тотығуының төмен тиімділігі митохондриялық тыныс алу үшін субстрат қолжетімділігін төмендетуге ықпал етеді. (9) Гликолиз тотығу фосфорлануында АТФ өндірісінің төмендеуінің компенсаторлық механизмі ретінде күшейтіледі. (10) ATP синтазасы ?m-ді сақтау үшін кері бағытта жұмыс істейді. Қызыл және көк сәйкесінше жоғары және төмен реттеуді білдіреді. Қораптар эксперименттік дәлелдемелердің болуын білдіреді. Кірістірілгенде потенциометриялық флуоресцентті зонд тетраметилродамин метил эфирі (TMRM; 2 нМ) арқылы көрнекіленген WT және Nrf25-KO кортикальды астроциттердің митохондрияларының кескіндері көрсетілген. Масштаб жолағы, 20 μм.
Генотиптер арасындағы ??м мен тыныс алудағы бұл айырмашылықтар митохондриялық тыныс алу үшін субстраттарды пайдалану жылдамдығымен көрінеді. Үшкарбон қышқылының (TCA) циклі үшін субстраттарды қолдану (малат/пируват, ол өз кезегінде I NADH кешенінің өндірісін арттырады) немесе метилсукцинат, II кешеннің субстраты екі WT-де де ??м-нің сатылы өсуін тудырады. және Keap1-KD нейрондары, бірақ өсу жылдамдығы Keap1-KD жасушаларында жоғары. Ең бастысы, бұл TCA циклінің субстраттарына жауаптың пішіндері екі генотиптің арасында әртүрлі, соның арқасында субстрат қосылған кезде Keap1-KD жасушаларында ?m-нің жылдам өсуі плато емес, тез төмендеумен жалғасады, бұл әдеттен тыс болуын болжайды. субстратты жылдам тұтыну. Бұл нәтижелер WT MEF-пен салыстырғанда Keap50-KO-да [U-70C1] глюкозаның 13 сағаттық импульсінен кейін байқалған малаттың, пируваттың және сукцинаттың әлдеқайда төмен (6-1%) деңгейлерімен тығыз сәйкес келеді. жасушалар [24]. Nrf2-KO нейрондарында тек пируват ?m арттыруға қабілетті, ал малат пен метилсукцинат жеңіл деполяризацияны тудырады. Nrf2 митохондриялық субстрат өндірісіне әсері Nrf2 митохондриялық функцияға әсер ететін негізгі механизм болып көрінеді. Митохондриялық NADH тотығу-тотықсыздану индексі (NADH I комплексінің тұтынуы мен TCA цикліндегі NADPH өндірісі арасындағы тепе-теңдік) Nrf2-KO жасушаларында олардың WT аналогтарымен салыстырғанда айтарлықтай төмен, сонымен қатар, бассейндердің регенерация жылдамдығы. IV комплексті тежегеннен кейін NADH және FADH2 (NaCN қолдану арқылы) мутантты жасушаларда баяуырақ.
Мысықтың миы мен бауырынан оқшауланған митохондрияларда I немесе II кешен үшін субстраттарды толықтыру Nrf2 белсендірілген кезде оттегін тұтыну жылдамдығын күштірек арттырады және Nrf2 бұзылған кезде тиімділігі азырақ [35]. Осылайша, малат WT салыстырғанда Keap1-KD оттегін тұтынудың жоғары жылдамдығын тудырады, бірақ Nrf2-KO митохондрияларында оның әсері әлсіз. Сол сияқты, ротенонның қатысуымен (комплекс I тежелген кезде) сукцинат WT-мен салыстырғанда Keap1-KD-де оттегі тұтынуды көбірек белсендіреді, ал Nrf2-KO митохондрияларында жауап азаяды. Сонымен қатар, Nrf2-KO негізгі нейрондық дақылдары мен тышқандар II кешенді ингибиторлары 3-нитропропион қышқылы мен малонаттың уыттылығына аса сезімтал, ал Nrf2 артық экспрессиялық астроциттердің интрастриатальды трансплантациясы қорғаныс болып табылады [48], [49]. Сол сияқты, Nrf2-KO тышқандары Nrf2-нің генетикалық немесе фармакологиялық активтенуі 1-метил-4-фенил-1-дегі 4-метил-1,2,3,6-фенилпиридиний ионының I комплексті тежегішінен туындаған нейроуыттылыққа қарсы қорғаныс әсеріне ие болады, Паркинсон ауруының 49-тетрагидропиридинді жануар үлгісі [50], [51], [52], [53], [54], [55], [56], [57], [58], [59], [60], [61], [XNUMX].
Тыныс алуды бақылау коэффициенті (RCR), 3-күйдің (ADP-стимулданған) 4-күйге қатынасы (ADP жоқ) Nrf2 болмаған кезде төмендейді, бірақ RCR Keap1-KD және WT митохондриялары арасында ұқсас [35] ]. RCR митохондриялық тыныс алу тізбегі белсенділігінің тотығу фосфорлануына қосылу дәрежесінің көрсеткіші болғандықтан, бұл нәтиже Keap1-KD митохондрияларындағы тыныс алудың жоғары жылдамдығы тотығу фосфорлануының ажырауымен байланысты емес екенін көрсетеді. Бұдан әрі ол Nrf2 белсендірілген кезде тотығу фосфорлануы тиімдірек болады деп болжайды. Keap1-KD митохондрияларындағы тыныс алудың жоғары жылдамдығы митохондриялық ROS өндірісінің жоғары деңгейіне сәйкес келеді [38], өйткені жоғары тыныс алу жылдамдығы электрондардың ағып кетуіне әкелуі мүмкін. Дегенмен, тотығу стрессі жағдайында ROS өндірісінің жоғарылауы митохондрияның ішкі мембранасының протон өткізгіштігін жоғарылататын және нәтижесінде супероксид өндірісін төмендететін 2-ші ақуыздың (UCP3) Nrf3-тәуелді транскрипциялық жоғарылауымен қарсы әрекет етеді [62]. Жақында липидтердің асқын тотығу өнімі 4-гидрокси-2-ноненал кардиомиоциттерде UCP2-тің Nrf3-тәуелді жоғарылауына делдал болатыны көрсетілді; бұл әсіресе ишемия-реперфузия кезіндегі тотығу стрессі жағдайында қорғаныс үшін маңызды болуы мүмкін [63].
Nrf2 тотығу фосфорлануының және АТФ синтезінің тиімділігіне әсер етеді
Nrf2-нің тыныс алуға, ми мен бауыр митохондрияларына әсерімен келісе отырып, Nrf2 тапшылығы тотығу фосфорлану тиімділігінің төмендеуіне әкеледі (АТФ синтезі үшін жұмсалатын ADP-тің оттегіге қатынасы бойынша бағаланады), ал Nrf2 белсендіруі (Keap1) -ҚД) кері әсер етеді [35]. WT-мен салыстырғанда, ATP деңгейлері Nrf2 конститутивті жоғарылауы бар жасушаларда айтарлықтай жоғары және Nrf2 құлаған кезде [64] немесе бұзылғанда [35] төмен. Сонымен қатар, тотығу фосфорлану (олигомицин) немесе гликолиз (йодасірке қышқылы) ингибиторларын қолдану Nrf2 жасушаларының ATP түзу жолын өзгертетінін анықтады. Осылайша, WT нейрондарында олигомицин АТФ толық төмендеуін тудырады және йодосірке қышқылы одан әрі әсер етпейді. Бір қызығы, Nrf2-KO жасушаларында олигомицин йодосірке қышқылымен баяу, бірақ толығымен таусылатын ATP деңгейін жоғарылатады, бұл Nrf2 болмаған кезде тотығу фосфорлану емес, гликолиз АТФ өндірісінің негізгі көзі болып табылатынын көрсетеді. Бір қызығы, Keap1-KD жасушаларында тотығу фосфорлану тиімділігінің жоғарылауына қарамастан, олигомицинді қосу ATP деңгейінің ~80% төмендеуіне әкеледі, ал йодосірке қышқылы одан әрі ~20% төмендеуіне әкеледі. Осылайша, Nrf2 тапшылығы немесе оның конститутивтік активтенуі тотығу фосфорлану үлесін азайтады және АТФ синтезіне гликолиздің үлесін арттырады. Бұл әсер әсіресе Nrf2 болмаған кезде айқын көрінеді және ??m ортадағы глюкозаның болуына [35] тәуелділігіне және гликолитикалық аралық өнімдердің (G-6-P, F-6-P) жоғарылауына сәйкес келеді. , дигидроксиацетонфосфат, пируват және лактат) Nrf2 [24] құлағаннан кейін.
Олигомицинмен F1F0-ATPase тежеуінен кейін АТФ деңгейінің жоғарылауы Nrf2 болмаған жағдайда F1F0-ATPase ATP синтаза емес, АТФ-аза ретінде қызмет ететінін көрсетеді, яғни ол кері бағытта жұмыс істейді. Белсенділіктің мұндай өзгеруі, ең алдымен, осы органелланың функционалдық тұтастығы үшін маңызды болып табылатын ?m-ді сақтау әрекеті үшін протондарды ішкі митохондриялық мембрана арқылы айдау қажеттілігін көрсетеді. F1F0-ATPase функциясының кері өзгеруі, сондай-ақ, Nrf2-KO жасушаларына олигомицинді енгізу кезінде байқалған митохондриялық деполяризациямен дәлелденеді, бұл олардың WT немесе Keap1 тапшылығы бар аналогтарында болатын гиперполяризациядан күрт айырмашылығы бар [35]. Тұтастай алғанда, Nrf2 тапшылығы жағдайында АТФ негізінен гликолизде өндірілетін сияқты, және бұл АТФ кейіннен F1F0-ATPase арқылы ішінара ??m ұстап тұру үшін пайдаланылады.
Nrf2 митохондриялық май қышқылдарының тотығуын жақсартады
Nrf2 тапшылығының ??m-ге әсері, әсіресе жасушалар глюкозасыз ортада инкубацияланғанда айқын көрінеді, ал ??m WT жасушаларымен салыстырғанда Nrf50-KO-да ~2% төмен [35]. Глюкоза жетіспеушілігі жағдайында митохондриялық май қышқылдарының тотығуы (ФАО) тыныс алу және тотығу фосфорлануы үшін субстраттардың негізгі жеткізушісі болып табылады, бұл Nrf2 ФАО-ға әсер етуі мүмкін екенін көрсетеді. Шынында да, ФАО-ның ұзақ тізбекті (C16:0) қаныққан май қышқылы пальмитин қышқылы үшін де, қысқа тізбекті (C6:0) гексоной қышқылы үшін де тиімділігі Keap1-KO MEFs және оқшауланған жүрек пен бауыр митохондрияларында олардың тиімділігіне қарағанда жоғары. WT аналогтары, ал Nrf2-KO жасушаларында және митохондрияларда төменірек [65]. Бұл әсерлер адамдар үшін де өте маңызды: шынында да, ФАО-ның TCA циклінің белсенділігімен жақсырақ интеграциялануын көрсететін метаболикалық өзгерістер классикалық Nrf2 сульфорафан активаторының прекурсоры глюкорафанинге бай диеталармен адамның араласуын зерттеуде орын алғаны туралы хабарланған. 66].
Митохондриялық ФАО-ның бірінші сатысында ?-көміртегінің pro-R сутегісі FAD кофакторын FADH2-ге дейін төмендететін гидрид ретінде кетеді, бұл өз кезегінде электрондарды тыныс алу тізбегіндегі убихинонға (UbQ) тасымалдап, сайып келгенде АТФ түзілуіне ықпал етеді. . Глюкозаның жоқтығында ФАО-ны пальмитойлкарнитинмен ынталандыру WT және Keap1-KO жасушаларында ATP деңгейінің күтілетін жоғарылауын тудырса, Keap1-KO жасушаларында ATP жоғарылауы жылдамырақ болса, бірдей емдеу Nrf2-KO-да АТФ өзгерістерін тудырмайды. MEFs [65]. Бұл эксперимент Nrf2 болмаған жағдайда ФАО басылатынын көрсетеді, сонымен қатар ол Nrf2 тапшылығы жағдайында АТФ деңгейінің төмендеуінің себептерінің бірі ретінде ФАО-ның басылуын көрсетеді [35], [64].
Атап айтқанда, Nrf293 дыбысы өшірілген адамның 2 Т жасушаларында CPT1 және CPT2 [67], митохондриялық ФАО жылдамдығын шектейтін фермент карнитин пальмитоилтрансферазаның (CPT) екі изоформасы төменірек экспрессияға ие. Келісім бойынша, Cpt1 мРНҚ деңгейлері WT тышқандарымен салыстырғанда Nrf2-KO бауырларында төмен [68]. CPT ұзын тізбекті майлы ацил-КоА ацил тобының кофермент А-дан l-карнитинге ауысуын катализдейді және осылайша цитоплазмадан митохондрияға ацилкарнитиннің импортына мүмкіндік береді. Бұл бүгінгі күнге дейін зерттелмегенімен, CPT1 экспрессиясына транскрипциялық әсерлерден басқа, Nrf2 оның негізгі аллостериялық ингибиторы малонил-КоА деңгейін бақылау арқылы осы ферменттің қызметіне де әсер етуі мүмкін. Бұл қазіргі уақытта түсініксіз механизм арқылы Nrf2 стероил КоА десатураза (SCD) [69] және цитрат-лиаза (CL) [69], [70] экспрессиясын теріс реттейді. Бір қызығы, SCD нокаут немесе тежелу фосфорланудың жоғарылауына және AMP-белсендірілген протеинкиназаның (AMPK) белсендірілуіне әкеледі [71], [72], [73] және Nrf2 болмаған жағдайда SCD деңгейлері деп болжауға болады. артады, өз кезегінде AMPK белсенділігін төмендетеді. Бұл Nrf2-KO тышқандарының [68] бауырында байқалған AMPK ақуыз деңгейінің төмендеуімен одан әрі күшейтілуі мүмкін, бұл Keap1-KD бауырларында хабарланған AMPK деңгейінің жоғарылауымен тығыз сәйкес келеді. тышқандар [74]. AMPK белсенділігінің төмендеуінің бір салдары оның ацетил-КоА карбоксилазасының (ACC) ингибиторлық фосфорлануының (Ser79 деңгейінде) әлсіреуі болып табылады [75], ол Nrf2 болмаған кезде одан әрі транскрипциялық түрде жоғары реттелуі мүмкін, өйткені ол Nrf2 белсендіруімен төмендейді [70] ]. ACC субстраты ацетил-КоА өндірісін арттыратын жоғары реттелетін CL экспрессиясымен үйлесетін жоғары ACC белсенділігі, сайып келгенде, ACC өнімінің, малонил-КоА деңгейін арттыруы мүмкін. Малонил-КоА жоғары деңгейлері CPT тежейді, осылайша май қышқылдарының митохондрияға тасымалдануын азайтады. Ақырында, Nrf2 плазма мен митохондриялық мембраналар арқылы май қышқылдарын импорттайтын транслоказа - CD36 [76] экспрессиясын оң реттейді. Осылайша, Nrf2 митохондриялық ФАО тиімділігіне әсер етуі мүмкін механизмдердің бірі митохондрияға ұзын тізбекті май қышқылдарының импортын реттеу болып табылады.
Тікелей транскрипциялық реттеуден басқа, Nrf2 жасушалық тотығу-тотықсыздану метаболизміне әсері арқылы митохондриялық ФАО тиімділігін өзгертуі мүмкін. Бұл әсіресе Nrf2 белсенділігі төмен немесе жоқ болғанда, жасушалық тотығу-тотықсыздану күйін тотыққан күйге ауыстыратын жағдайларда маңызды болуы мүмкін. Шынында да, бірнеше ФАО ферменттері тотығу-тотықсыздану өзгерістеріне сезімтал екені анықталды. Осындай ферменттердің бірі - өте ұзын тізбекті ацил-КоА дегидрогеназа (VLCAD), ол адам тіндеріндегі пальмитойл-КоА дегидрогенизация белсенділігіне 80%-дан астам үлес қосады [77]. Бір қызығы, Hurd et al. [78] VLCAD құрамында оқшауланған егеуқұйрық жүрек митохондрияларына H2O2 әсер еткенде тотығу-тотықсыздану күйін айтарлықтай өзгертетін цистеин қалдықтары бар екенін көрсетті. Сонымен қатар, Cys238-де тышқан бауырының VLCAD S-нитрозилизациясы ферменттің каталитикалық тиімділігін жақсартады [79] және сол цистеиннің тотығуы кері әсер етуі мүмкін, сайып келгенде, митохондриялық ФАО тиімділігін төмендетеді. Сондықтан, VLCAD экспрессия деңгейлері WT, Nrf2-KO немесе Keap1-KO MEF [65]-де айтарлықтай ерекшеленбесе де, VLCAD ферментінің белсенділігі жоғары деңгейлерге байланысты Nrf2 болмаған кезде төмен болуы мүмкін. ROS.
Осы тұжырымдардың барлығына сүйене отырып, мынаны ұсынуға болады (3-сурет): Nrf2 болмаған кезде, ME1, IDH1, G6PD және PGD экспрессиясының төмендеуіне байланысты NADPH деңгейлері төмен. Азайған глутатион деңгейі оның биосинтезі мен регенерациясына қатысатын ферменттердің экспрессиясының төмендеуіне және тотыққанды глутатионның тотықсызданған түріне айналдыру үшін қажет NADPH деңгейінің төмендеуіне байланысты да төмен. ME1-тің төмен экспрессиясы митохондрияға түсетін пируват пулын азайтады, гликолиз пируваттың негізгі көзіне айналады. NADH генерациясы баяу, бұл I кешенінің белсенділігінің бұзылуына және митохондриялық ROS өндірісінің жоғарылауына әкеледі. FAD-ның FADH2-ге дейін төмендеуі де баяуырақ, кем дегенде ішінара май қышқылдарының тиімді тотығуына байланысты, FADH2-ден UbQ-ға және III комплекске электрон ағынын бұзады. UbQH2 сукцинатдегидрогеназаның активаторы болғандықтан [80], оның түзілуін бәсеңдету сукцинатдегидрогеназа ферментінің белсенділігін төмендетуі мүмкін. Супероксид пен сутегі асқын тотығының жоғарылауы кешенді II белсенділікті одан әрі тежей алады [81]. Май қышқылдарының тотығуының төмен тиімділігі митохондриялық тыныс алу және тотығу фосфорлану кезінде АТФ өндірісі үшін субстрат қолжетімділігінің төмендеуіне ықпал етеді. Компенсаторлық механизм ретінде гликолиз күшейеді. ATP синтазасы ?m-ді сақтау әрекетінде АТФаза ретінде керісінше жұмыс істейді.
Nrf2 және митохондриялық биогенез
WT-мен салыстырғанда, Nrf2-KO тышқандарының бауырында митохондриялық мазмұн төмен (митохондрияның ядролық ДНҚ-ға қатынасымен анықталады) екені хабарланды; бұл WT және Nrf24-KO тышқандарының екеуінде де 2 сағаттық жылдамдықпен одан әрі төмендейді; керісінше, қалыпты азықтандыру жағдайында WT-ден айырмашылығы болмаса да, жоғары Nrf2 белсенділігі бар тышқандардағы митохондриялық мазмұн ораза ұстауға әсер етпейді [82]. Бір қызығы, Nrf2 активаторымен (R)-?-липой қышқылымен толықтыру [83], [84], [85] 3T3-L1 адипоциттеріндегі митохондриялық биогенезге ықпал етеді [86]. Ядролық транскрипциялық реттегіштердің екі класы митохондриялық биогенезде маңызды рөл атқарады. Бірінші класқа транскрипция факторлары жатады, мысалы, бес респираторлық кешеннің суббірліктерін кодтайтын гендердің экспрессиясын бақылайтын ядролық респираторлық факторлар11 және 2, митохондриялық трансляциялық компоненттер және митохондриялық матрицаға локализацияланған гем биосинтетикалық ферменттер [88]. Piantadosi және т.б. [89] 2-ядролық респираторлық фактордың Nrf1-тәуелді транскрипциялық жоғарылауы митохондриялық биогенезге ықпал ететінін және кардиоуытты антрациклин химиотерапевтік агенті доксорубициннің цитотоксикалық әсерінен қорғайтынын көрсетті. Керісінше, Чжан және т.б. [82] Nrf2-нің генетикалық активтенуі тышқан бауырындағы 1-ядролық респираторлық фактордың базальды мРНҚ экспрессиясына әсер етпейтінін хабарлады.
Митохондриялық биогенезде маңызды функциялары бар ядролық транскрипциялық реттеушілердің екінші класы транскрипциялық коактиваторлар болып табылады, мысалы пероксисома пролифераторымен белсендірілген рецептор? коактиваторлар (PGC)1? және 1?, олар транскрипция факторларымен, базальды транскрипциялық және РНҚ-қосқыш аппаратымен және гистонды өзгертетін ферменттермен әрекеттеседі [88], [90], [91]. PGC1 коактиваторлар тобының экспрессиясына көптеген қоршаған орта сигналдары әсер етеді. Nrf2 активатор сульфорафанмен адам фибробласттарын өңдеу митохондриялық массаның ұлғаюын және PGC1 индукциясын тудырады? және PGC1? [92], дегенмен Nrf2-ге ықтимал тәуелділік бұл зерттеуде зерттелмеген. Дегенмен, Nrf2 Keap1 генінің гипоморфты нокдаунымен (db/db:Keap1flox/?:Nrf2+/+) белсендірілген немесе бұзылған (db/db:Keap1flox/?:Nrf2?/?) диабеттік тышқандарда бауыр PGC1 төмен ме? өрнек деңгейлері бақылау жануарларына қарағанда (db/db:Keap1flox/+:Nrf2+/+) [93]. PGC1 үшін мРНҚ деңгейлерінде айырмашылық жоқ па? WT немесе Nrf2-KO болатын диабеттік емес тышқандардың бауырында байқалады, ал бұл деңгейлер Nrf2 шамадан тыс экспрессияланатын (Keap1-KD және бауырға тән Keap1-KO) жануарларда төмен [82]. Айта кетейік, 24 сағаттық ораза PGC1 деңгейін жоғарылатады ма? mRNA барлық генотипті тышқандардың бауырында, бірақ WT немесе Nrf2 шамадан тыс экспрессивті тышқандармен салыстырғанда Nrf2-KO бауырларында ұлғайған. WT-мен салыстырғанда, септикалық инфекцияны немесе инфекция салдарынан жедел өкпе жарақатын бастан кешірген Nrf2-KO тышқандары 1 ядролық тыныс алу факторының және PGC1-нің әлсіреген транскрипциялық жоғарылауын көрсетеді? [94], [95]. Бұл бақылаулар бірге Nrf2-нің ядролық тыныс алу факторы 1 және PGC1 деңгейін ұстап тұрудағы рөлін көрсетеді? күрделі және күйзеліс жағдайында ең көрнекті болады.
Митохондриялық ақуыздарды кодтайтын гендердің экспрессиясынан басқа, митохондрия биогенезі нуклеотидтердің синтезін қажет етеді. Nrf2 генетикалық белсендіру пентозофосфат жолын және фолий мен глутамин метаболизмін, әсіресе тез таралатын жасушаларда жоғары реттеу арқылы пурин биосинтезін күшейтеді (Cурет 2) [24]. Митохондриялық серин/треонин протеинкиназасы PTEN-индукцияланған болжамды киназа 1 (PINK1) үшін жетіспейтін мутантты дрозофиланың транскриптомының талдауы митохондриялық дисфункцияның нуклеотидтер метаболизміне әсер ететін гендердің транскрипциялық жоғарылауына әкелетінін көрсетті. PINK96 тапшылығының нейротоксикалық салдарынан қорғау механизмін білдіреді. Nrf1 де жаңа пуриндік нуклеотидтің биосинтетикалық жолына кіруді катализдейтін фосфорибозилпирофосфат амидотрансферазасының (PPAT) және митохондриялық метилентетрагидрофолатдегидрогеназа 2 (MTHFD2) экспрессиясын реттейді (2-сурет). Соңғысы дегидрогеназа және циклогидролаза белсенділігі бар екі функционалды фермент болып табылады, ол тез өсіп келе жатқан жасушаларда пурин биосинтезі үшін бір көміртекті бірлік көздері ретінде глицин мен форматты қамтамасыз етуде маңызды [2]. Сондықтан Nrf97 белсендіру қорғаныш болуы мүмкін және PINK2 тапшылығында митохондриялық дисфункцияны қалпына келтіруі мүмкін. Шынында да, сульфорафан немесе тритерпеноид RTA-1 арқылы Nrf2-нің фармакологиялық активтенуі ?m қалпына келтіреді және PINK408 жетіспейтін жасушаларды дофамин уыттылығынан қорғайды [1]. Негізгі механизмдер күрделі болып көрінгенімен, бірге бұл нәтижелер Nrf98 белсенділігі транскрипцияның сыни факторлары мен коактиваторларының экспрессиялық деңгейлеріне әсер ету, сондай-ақ нуклеотидтер биосинтезін күшейту арқылы митохондрия биогенезіне әсер етуі мүмкін екенін көрсетеді.
Nrf2 және митохондриялық тұтастық
Тікелей дәлелдер әрқашан қол жетімді болмаса да, Nrf2 митохондриялық тұтастық үшін, әсіресе тотығу стрессі жағдайында маңызды екендігінің күшті белгілері бар. Nrf2 белсендіргіш сульфорафанның бір реттік дозасын енгізген егеуқұйрықтардың миы мен бауырынан бөлінген митохондриялар тотықтырғыш терт-бутилгидропероксид тудыратын митохондриялық өткізгіштіктің өту тесігінің (mPTP) ашылуына төзімді [99], [100]. mPTP, митохондриялық ішкі мембрананың массасы 1500 Да дейінгі молекулалар үшін өткізгіштігін қамтамасыз ететін кешен, жақында F0F1-ATP синтазасының димерлерінен түзілетіні анықталды [101]. Сульфорафан арқылы mPTP ашылуына төзімділік антиоксиданттық қорғаныстың жоғарылауымен корреляцияланады және митохондриялық GSH, глутатион пероксидаза 1, алма ферменті 3 және тиоредоксин 2 деңгейлері сульфорафанмен өңделген жануарлардан бөлінген митохондриялық фракцияларда жоғары реттеледі.
Митохондриялық ақуыздың зақымдалуы және электрофильді липидтердің асқын тотығу өнімі 4-гидрокси-2-ноненалдан туындаған тыныс алудың бұзылуы сульфорафанмен өңделген тышқандардың церебральды қыртысынан бөлінген митохондрияларда әлсірейді [102]. Егеуқұйрықтардың бүйрек эпителий жасушаларында және бүйректе сульфорафан цисплатин және гентамицин тудыратын уыттылықтан және ?м жоғалтудан қорғайды [103], [104]. Тотықтырғыштар панелінен (супероксид, сутегі асқын тотығы, пероксинитрит) және электрофилдерден (4-гидрокси-2-ноненал және акролеин) қорғаныс және егеуқұйрық аортасының тегіс бұлшықет жасушаларын сульфорафанмен өңдеген кезде де митохондриялық антиоксиданттық қорғаныстың жоғарылауы байқалды [105] ]. Жақында контрастпен индукцияланған бүйректің жедел зақымдалу үлгісінде аяқ-қолдың ишемиялық алдын-алуының GSK2 тежелуінің нәтижесінде Nrf3 белсендіруі арқылы mPTP ашылуын тежеуді және митохондриялық ісінуді қоса алғанда, қорғаныс әсерлері бар екені көрсетілді? [106].
Митофагия, яғни дисфункционалды митохондрияларды аутофагосомалар іріктеп жұтып, лизосомаларға жасуша ыдырауы және қайта өңдеуі үшін жеткізілетін процесс, митохондриялық гомеостаз үшін маңызды [107], [108]. Nrf2 мен митофагия арасында себеп-салдарлық байланыс анықталмағанымен, транскрипция факторы митохондриялық сапаны бақылауда митофагияда рөл атқара отырып, маңызды болуы мүмкін екендігі туралы дәлелдер бар. Бұл әсіресе тотығу стрессі жағдайында айқын болуы мүмкін. Осылайша, сепсис үлгісінде WT тышқандарымен салыстырғанда Nrf1-KO-да MAP3 жеңіл тізбегі 3-II (LC62-II) және жүк ақуызы p24 деңгейінің жоғарылауы Nrf2-KO-да басылады [109] . Митофагияның шағын молекулалы индукторы (p62-митофагия индукторы, PMI деп аталады) жақында ашылды; бұл 1,4-дифенил-1,2,3-триазол қосылысы бастапқыда транскрипция факторының Keap2-мен әрекеттесуін бұзатын Nrf1 активаторы ретінде жасалған [110]. Nrf2 генетикалық жоғары реттелетін жасушаларға ұқсас (Keap1-KD немесе Keap1-KO), PMI әсеріне ұшыраған жасушалардың демалу деңгейі жоғарырақ болады. Маңыздысы, WT жасушаларының PMI өңдеуінен кейін байқалатын митохондриялық LC3 локализациясының жоғарылауы Nrf2-KO жасушаларында болмайды, бұл Nrf2 қатысуын болжайды.
Соңғысы, бауыр бөлімдерінің ультрақұрылымдық талдауы 2 апта бойы майы жоғары диетаны тамақтандырған тышқандарда WT емес, Nrf24-KO гепатоциттерінде кристалдары төмендеген және жарғақшалары бұзылған ісінген митохондриялардың болуын анықтады; атап айтқанда, бұл бауырлар тотығу стрессі мен қабынудың айқын дәлелдерін көрсетеді [68]. Nrf2 тотығу және қабыну стресс жағдайында митохондриялық тұтастықты сақтауда маңызды рөл атқарады деп қорытынды жасауға болады.
Сульфорафан және оның қатерлі ісікке, өлімге, қартаюға, ми мен мінез-құлыққа, жүрек ауруына және басқаларына әсері
Изотиоцианаттар сіздің диетаңызға түсетін ең маңызды өсімдік қосылыстарының бірі болып табылады. Бұл бейнеде мен олар үшін жасаған ең жан-жақты жағдайды жасаймын. Қысқаша назар аударыңыз ба? Төмендегі уақыт нүктелерінің бірін таңдап, сүйікті тақырыпқа өтіңіз. Толық уақыт шкаласы.
Негізгі бөлімдер:
00: 01: 14 - қатерлі ісік және өлім
00: 19: 04 - қартаю
00: 26: 30 - Ми және мінез
00: 38: 06 - Соңғы қорытынды
00: 40: 27 - Доза
Толық уақыт шкаласы:
00: 00: 34 - бейнефильмнің басты тақырыбы - сульффоран енгізу.
00: 01: 14 - Өсімдік тұтыну және барлық өлім-жітімде қысқарту.
00: 02: 12 - простата қатерлі ісігі.
00: 02: 23 - Мочевина рагы қатер.
00: 02: 34 - темекі шегудегі қатерлі ісік.
00: 02: 48 - Сүт безі қатерлігі қатері.
00: 03: 13 - Гипотетикалық: егер сізде рак бар болса? (интервенционалдық)
00: 03: 35 - қатерлі ісік пен өлім қауымдасқан деректерді басқаратын шынайы механизм.
00: 04: 38 - сульфорфан және қатерлі ісік.
00: 05: 32 - егеуқұйрықтардағы ісік ісігін дамытуға арналған брокколи өсімдік сығындысының күшті әсерін көрсететін жануарлы дәлел.
00: 26: 30 - мидың және мінез-құлық бөлімінің басы.
00: 27: 18 - аутизмге арналған брокколи өсімдік сығындысының әсері.
00: 27: 48 - глюкурофенияларға арналған глюкурофанның әсері.
00: 28: 17 - Депрессияның талқылануын бастау (ықтимал механизм мен зерттеулер).
00: 31: 21 - Стресс-индуцирленген депрессияның 10 түрлі модельдерін қолданып тышқандарды зерттеу, сульфарфетан сияқты флоксин (prozac) сияқты тиімді.
00: 32: 00 - зерттеу тышқандарда глюкарофаниннің тікелей ішуін көрсетеді, социологиялық жеңілдетудің стресс-моделінен депрессияны болдырмау үшін де тиімді.
00: 33: 01 - нейродегенерация бөлімінің басы.
00: 33: 30 - сульфорфан және Альцгеймер ауруы.
00: 33: 44 - Сульфорфан және Паркинсон ауруы.
00: 33: 51 - Сульфорфан және Хантингтон ауруы.
00: 34: 13 - Сульфорфан жылу әсерін арттырады.
00: 34: 43 - бас миының зақымдану секциясының басы.
00: 35: 01 - Туберкулезге қарсы тестілеуден кейін бірден сульфорфан препараты (тышқанның зерттеуі).
00: 35: 55 - сульфорфан және нейрондық пластика.
00: 36: 32 - Сулфорафан тышқандар типіндегі II типті қант диабеті моделін үйренуді жақсартады.
00: 37: 19 - сульфорфан және дученне бұлшықет дистрофиясы.
00: 38: 06 - Кешіктіріп бейнелеу: өлім және қатерлі ісік, ДНҚ зақымдануы, тотығу стресті және қабыну, бензолдың экскрециясы, жүрек-қан тамырлары аурулары, II типті диабет, миға әсер ету (депрессия, аутизм, шизофрения, нейрогенерация), NRF2 жолдары.
00: 40: 27 - брокколи өскіндер немесе сульффоранның дозасын анықтау туралы ойлар.
00: 41: 01 - Үйде өсіруге арналған анекдоттар.
00: 43: 14 - Пісіру температуралары және сульфарфан белсенділігі туралы.
00: 43: 45 - Глюкорафаниннен сульффоранның Gut бактерияларына айналуы.
00: 44: 24 - Көкөністерден белсенді мирозиназмен үйлескен кезде қоспалар жақсы жұмыс істейді.
00: 44: 56 - пісіру әдісі және крест тәрізді көкөністер.
Nrf2 - адам ағзасының жасушалық антиоксиданттық қорғаныс жүйесінде маңызды рөл атқаратын транскрипция факторы. Антиоксидантты жауап беретін элемент немесе ARE гендердің реттеуші механизмі болып табылады. Көптеген зерттеу зерттеулері Nrf2 немесе NF-E2 байланысты фактор 2 жасушалардың бірнеше түрлерінде ARE басқаратын гендердің кең ауқымын реттейтінін көрсетті. Nrf2 сонымен қатар жасушаны қорғауда және канцерогенділікке қарсы маңызды рөл атқаратыны анықталды, бұл Nrf2 тотығу стрессінен туындаған нейродегенеративті ауруларды және қатерлі ісіктерді басқаруда тиімді ем болуы мүмкін екенін көрсетеді. Доктор Алекс Хименес DC, CCST Insight
Қорытынды ескертулер
Көптеген сұрақтар әлі де ашық болса да, қолда бар эксперименттік дәлелдер Nrf2 митохондриялық гомеостазды және құрылымдық тұтастықты қолдаудағы маңызды ойыншы екенін анық көрсетеді. Бұл рөл Nrf2-делдалдық цитопротекторлық реакцияларды реттеу қабілеті жасуша мен ағзаның жалпы денсаулығына және өмір сүруіне әсер еткенде тотығу, электрофильді және қабыну стресс жағдайында өте маңызды болады. Митохондриялық функциядағы Nrf2 рөлі осы транскрипция факторымен ұйымдастырылған кең цитопротекторлық механизмдердің басқа қабатын білдіреді. Көптеген адамның патологиялық жағдайларында тотығу стрессі, қабыну және митохондриялық дисфункция олардың патогенезінің маңызды құрамдас бөліктері болғандықтан, Nrf2-нің фармакологиялық белсендірілуі аурудың алдын алу мен емдеуге уәде береді. Nrf2 митохондриялық функцияға әсер ететін нақты механизмдерді жан-жақты түсіну болашақ клиникалық зерттеулерді ұтымды жобалау үшін өте маңызды және терапевтік тиімділікті бақылау үшін жаңа биомаркерлерді ұсына алады.
Жоғарыдағы мақаланың мақсаты митохондриялық функциядағы Nrf2-нің пайда болатын рөлін талқылау және көрсету болды. Nrf2 немесе ядролық фактор эритроид 2-байланысты фактор, тотығу стрессіне ықпал ете алатын, жасушалық функцияға әсер ететін және уыттылықтың, созылмалы аурулардың және тіпті қатерлі ісіктің дамуына әкелетін тотықтырғыштарға жасушалық төзімділіктің дамып келе жатқан реттеушісі. Адам ағзасындағы тотықтырғыштардың өндірісі жасушалардың бөлінуін, қабынуды, иммундық функцияны, аутофагияны және стресске жауап беруді қоса алғанда, әртүрлі мақсаттарға қызмет етуі мүмкін болса да, денсаулыққа қатысты мәселелердің алдын алу үшін олардың шамадан тыс өндірілуін бақылау өте маңызды. Біздің ақпаратымыздың көлемі хиропротикалық және омыртқаның денсаулығына қатысты мәселелермен шектеледі. Тақырыпты талқылау үшін доктор Хименеске хабарласыңыз немесе бізге телефон арқылы хабарласыңыз915-850-0900 .
Арқа ауруыWorldwide бұл мүгедектіктің ең көп таралған себептерінің бірі және бүкіл әлемде жұмыссыз қалған күндер. Арқадағы ауырсыну тек жоғарғы тыныс жолдарының инфекцияларымен санасқан дәрігерлерге жүгінудің екінші жалпы себебі болып табылады. Халықтың шамамен 80 пайызы өмір бойы кем дегенде бір рет арқа ауруы болады. Омыртқа - бұл басқа жұмсақ тіндермен қатар сүйектерден, буындардан, байламдардан және бұлшықеттерден тұратын күрделі құрылым. Осыған байланысты жарақаттар және / немесе ауырлататын жағдайлар, мысалықатты дискілер, ақыр соңында арқадағы ауырсыну белгілеріне әкелуі мүмкін. Спорттық жарақаттар немесе көлік апаты жарақаттары көбінесе арқадағы ауырсынудың жиі себебі болып табылады, бірақ кейде ең қарапайым қозғалыстар ауыр нәтижелерге әкелуі мүмкін. Бақытымызға орай, хиропрактикалық күтім сияқты емдеудің балама нұсқалары омыртқаны түзетулер мен қолмен манипуляцияларды қолдану арқылы арқадағы ауырсынуды жеңілдетуге көмектеседі, сайып келгенде, ауырсынуды жеңілдетеді.
Nrf2 Альцгеймер ауруы сияқты тотығу стресінің жоғарылауымен байланысты денсаулық мәселелерінің әсерінен адам ағзасын қорғайтын антиоксиданттық және детоксикациялаушы ферменттер мен гендер тобының белсендірілуін қолдайды. Нейродегенеративті аурулардың белгілерін басқаруға көмектесетін Nrf2 жолын белсендіру үшін әртүрлі табиғи заттар көрсетілді. Төмендегі мақаланың мақсаты - созылмалы қабынудан туындаған Nrf2 негізгі рөлін талқылау.
дерексіз
Қабыну канцерогенезде маңызды рөл атқара отырып, көптеген созылмалы аурулар мен асқынулардың ең жиі кездесетін белгісі болып табылады. Бірнеше зерттеулер Nrf2 қабыну жасушаларын тартуды ұйымдастыру және антиоксиданттық жауап элементі (ARE) арқылы ген экспрессиясын реттеу арқылы қабынуға қарсы процеске ықпал ететінін көрсетті. Keap1 (Келч тәрізді ECH-ассоциацияланған ақуыз)/Nrf2 (NF-E2 p45-байланысты фактор 2)/ARE сигналдық жолы негізінен қабынуға қарсы ген экспрессиясын реттейді және қабынудың дамуын тежейді. Сондықтан жаңа Nrf2-тәуелді қабынуға қарсы фитохимиялық заттарды анықтау дәрілік заттарды табудың негізгі нүктесі болды. Бұл шолуда біз Keap1/Nrf2/ARE сигналдық жолының мүшелерін және оның төменгі гендерін, осы жолдың қабыну ауруларының жануарлар үлгілеріне әсерін және NF-?B жолымен өзара байланысын талқылаймыз. Сонымен қатар, біз Nrf3 арқылы NLRP2 қабынуының реттелуін талқылаймыз. Бұдан басқа, Nrf2/ARE сигналдық жолына делдал болатын қабынуға қарсы фитохимиялық заттардың және басқалардың дамуының ағымдағы сценарийін қорытындылаймыз.
Қабыну – тіндер қоздырғыштар, зақымданулар немесе тітіркендіргіштер сияқты зиянды тітіркендіргіштермен жұқтырылған немесе зақымдалған кезде пайда болатын күрделі процесс. Бұл қорғаныс реакциясына иммундық жасушалар, қан тамырлары және молекулалық медиаторлар қатысады [1]. Қабыну сонымен қатар негізінен физикалық, химиялық, биологиялық және психологиялық факторлардың әсерінен туындайтын әртүрлі ауру күйлерімен байланысты патологиялық құбылыс болып табылады. Қабынудың мақсаты - жасушалық зақымның себептерін шектеу және жою, некротикалық жасушалар мен тіндерді тазарту және/немесе сіңіру және тіндердің қалпына келуін бастау. Қабынудың екі түрлі түрін ажыратады: жедел және созылмалы. Жедел қабыну өздігінен жойылады және иесіне пайдалы, бірақ ұзаққа созылған созылмалы қабыну көптеген созылмалы аурулар мен асқынулардың жалпы белгісі болып табылады. Моноциттер, макрофагтар, лимфоциттер және плазмалық жасушалар сияқты көптеген мононуклеарлы иммундық жасушалармен тікелей инфильтрация, сондай-ақ қабыну цитокиндерін өндіру созылмалы қабынуға әкеледі. Созылмалы қабыну канцерогенезде маңызды рөл атқаратыны белгілі [2]. Жалпы алғанда, қалыпты қабыну процесінде про- және қабынуға қарсы сигнал беру жолдары өзара әрекеттеседі.
Патологиялық қабыну процесінде алдымен мастикалық жасушалар, моноциттер, макрофагтар, лимфоциттер және басқа иммундық жасушалар белсендіріледі. Содан кейін жасушалар жарақат орнына жиналады, нәтижесінде макромолекулаларды, соның ішінде ДНҚ-ны зақымдайтын реактивті оттегі түрлері (ROS) пайда болады. Сонымен қатар, бұл қабыну жасушалары цитокиндер, хемокиндер және простагландиндер сияқты қабыну медиаторларының көп мөлшерін шығарады. Бұл медиаторлар одан әрі макрофагтарды қабынудың локализацияланған орындарына тартады және қабынумен байланысты көптеген сигнал беру каскадтарын және транскрипция факторларын тікелей белсендіреді. Қабынудың классикалық жолын дамытуға NF-?B (ядролық фактор каппа В), MAPK (митогенмен белсендірілген протеинкиназа) және JAK (янус киназа)-STAT (сигнал түрлендіргіштері және транскрипцияның активаторлары) сигналдық жолдары қатысады. [3], [4], [5]. Алдыңғы зерттеулер Nrf2 транскрипция факторы (NF-E2 p45 байланысты фактор 2) NADPH, NAD(P)H хиноноксидоредуктаза 1, глутатион пероксидаза, ферритин, гемоксигеназа-1HO (H1O) қоса, II фазаны детоксикациялаушы ферменттердің экспрессиясын реттейтінін анықтады. -6) және қабынуға қарсы әсері арқылы жасушаларды әртүрлі жарақаттардан қорғайтын антиоксиданттық гендер, осылайша аурудың ағымына әсер етеді [7], [8], [XNUMX].
Осы тамаша нәтижелерді ескере отырып, соңғы жылдары сигналдық жолдар арқылы қабыну ауруларына арналған мақсатты терапиялық препараттарды жасау үлкен қызығушылық тудырды. Бұл шолуда біз Keap1 (Келч тәрізді ECH байланысты протеин)/Nrf2 (NF-E2 p45-қа қатысты 2 фактор)/ARE (антиоксиданттық жауап элементі) қабынудағы сигналдық жолдағы зерттеулерді қорытындылаймыз.
Nrf2 құрылымы мен регламенті
Keap1-тәуелді Nrf2 ережесі
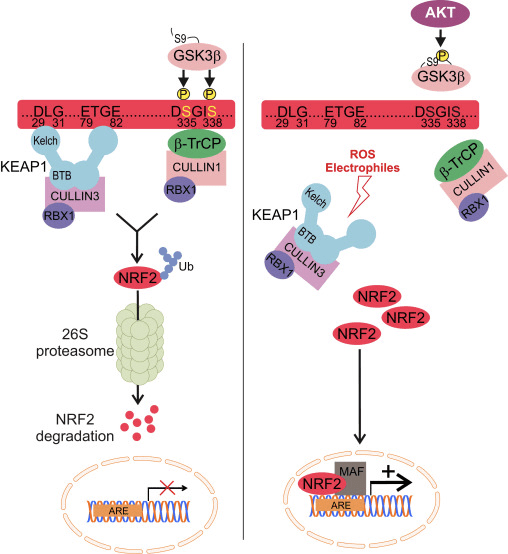
Nrf2 Cap �n� Collar (CNC) қосалқы отбасына жатады және жеті функционалды доменнен тұрады, Neh (Nrf2-ECH гомологиясы) 1 және Neh7 [9], [10]. Neh1 - Nrf2-ге кішкентай бұлшықет-апоневротикалық фибросаркома (Maf) протеині, ДНҚ және басқа транскрипция серіктестерімен гетеродимерленуге, сондай-ақ UbcM2 ubiquitin-конъюгациялаушы ферментімен ядролық кешен құруға мүмкіндік беретін CNC-bZIP домені [11], [12]. Neh2 құрамында DLG және ETGE деп аталатын екі маңызды мотив бар, олар Nrf2 мен оның теріс реттегіші Keap1 арасындағы өзара әрекеттесу үшін маңызды [13], [14].
Keap1 - қалыпты жағдайларда убиквитинация және протеазомалық деградация арқылы Nrf3 транскрипциялық белсенділігін тежейтін куллин негізіндегі E2 убиквитин лигазасының субстрат адаптері [15], [16], [17]. Keap1 гомодимерінің KELCH домендері цитозолдағы Nrf2-Neh2 доменінің DLG және ETGE мотивтерімен байланысады, мұнда ETGE жоғары жақындығы бар топса, ал DLG ілмек ретінде әрекет етеді [18]. Тотығу стрессі кезінде немесе Nrf2 активаторларының әсерінен Nrf2 Keap1 цистеинінің қалдықтарының тиолды модификациясына байланысты Keap1 байланысуынан ажырайды, бұл Nrf2 убиквитинациясын және протеазомалық деградацияны болдырмайды [19]. Содан кейін Nrf2 ядроға ауысады, кішкентай Maf ақуыздарымен гетеродимеризацияланады және гендердің ARE батареясын трансактивті етеді (1А-сурет). Neh3-тің карбокси-терминалы CHD6 (хромо-АТФаза/геликаза ДНҚ байланыстыратын ақуыз) деп аталатын транскрипцияның коактиваторымен әрекеттесу арқылы трансактивті домен ретінде әрекет етеді [20]. Neh4 және Neh5 сонымен қатар трансактивация домендері ретінде әрекет етеді, бірақ CBP (cAMP-жауап-элементті байланыстыратын ақуызды байланыстыратын ақуыз) деп аталатын басқа транскрипциялық коактиватормен байланысады [21]. Сонымен қатар, Neh4 және Neh5 RAC3/AIB1/SRC-3 ядролық кофакторымен өзара әрекеттеседі, бұл Nrf2 мақсатты ARE генінің экспрессиясының жоғарылауына әкеледі [22]. Neh5 Nrf2 реттеуі мен жасушалық локализациясы үшін өте маңызды болып табылатын тотықсыздандырғыш-сезімтал ядролық-экспорттық сигналға ие [23].
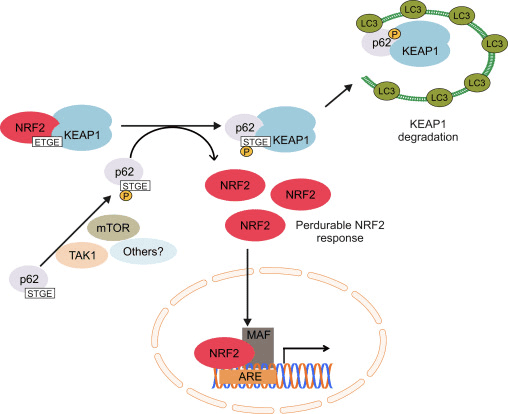
1-сурет Nrf1-нің Keap2-тәуелді және тәуелсіз реттелуі. (A) Базальды жағдайларда Nrf2 Keap1-мен екі мотивпен (ETGE және DLG) секвестрленеді, бұл CUL3-делдалдық убиквитинацияға, содан кейін протеазоманың деградациясына әкеледі. Тотығу стрессі кезінде Nrf2 Keap1-ден диссоциацияланады, ядроға ауысады және ARE-гендік батареяны белсендіреді. (B) GSK3 Nrf2 фосфорлайды және бұл CUL2-делдалдық убиквитинация және кейінгі протеазоманың ыдырауы үшін ?-TrCP арқылы Nrf1 танылуын жеңілдетеді. (C) p62 Keap1-мен секвестрленеді, бұл оның автофагиялық деградациясына, Nrf2 босатылуына және Nrf2 сигналының жоғарылауына әкеледі.
Keap1-Тәуелсіз Nrf2 ережесі
Пайда болған дәлелдер Keap2-ге тәуелсіз Nrf1 реттеуінің жаңа механизмін ашты. Nrf6 серинге бай Neh2 домені оның екі мотивімен (DSGIS және DSAPGS) ?-трансдуцинді қайталанатын ақуызға (?-TrCP) байланысу арқылы осы реттеуде шешуші рөл атқарады [24]. ?-TrCP - Skp1�Cul1�Rbx1/Roc1 убиквитин лигаза кешені үшін субстрат рецепторы, ол Nrf2 убиквитинациясы мен протеазомалық деградацияға бағытталған. Гликоген синтаза киназа-3 - Keap1-тәуелсіз Nrf2 тұрақтандыруға және реттеуге қатысатын маңызды ақуыз; ол Nrf2-ні ?-TrCP арқылы тануды және одан кейінгі ақуыз ыдырауын жеңілдету үшін Neh6 доменіндегі Nrf2-ні фосфорлайды [25] (1В-сурет).
Басқа Nrf2 реттегіштері
Басқа дәлелдер желісі p62-тәуелді Nrf2 активациясының канондық емес жолын ашты, онда p62 Keap1-ді аутофагиялық деградацияға секвестрлейді, бұл Nrf2-нің тұрақтануына және Nrf2-тәуелді гендердің трансактивациясына әкеледі [26], [27], [ 28], [29] (1С-сурет).
Жинақталған дәлелдер бірнеше miRNA Nrf2 белсенділігін реттеуде маңызды рөл атқаратынын көрсетеді [30]. Сангокоя және т.б. [31] miR-144 лимфобласт K2 жасушалық желісіндегі, адамның бастапқы эритроидты ізашар жасушаларында және орақ-жасушалы ретикулоциттердегі Nrf562 белсенділігін тікелей төмендететінін көрсетті. Адамның кеуде эпителий жасушаларында тағы бір қызықты зерттеу miR-28 Keap2-ге тәуелсіз механизм [1] арқылы Nrf32 тежейтінін көрсетті. Сол сияқты, miR-153, miR-27a, miR-142-5p және miR144 сияқты miRNAs нейрондық SH-SY2Y жасушалық желісіндегі Nrf5 экспрессиясын төмендетеді [33]. Сингх және т.б. [34] miR-93-тің эктопиялық экспрессиясы сүт бездерінің канцерогенезінің 2?-эстрадиол (E17)-индукцияланған егеуқұйрық үлгісінде Nrf2 реттелетін гендердің экспрессиясын төмендететінін көрсетті.
Біздің зертханада жақында ашылған жаңалық ретиноикалық X рецепторының альфасы (RXR?) деп аталатын Nrf2 эндогенді ингибиторын анықтады. RXR? ядролық рецептор болып табылады, Nrf7 Neh2 доменімен (амин қышқылдарының қалдықтары 209-316) ДНҚ байланыстыру домені (DBD) арқылы әрекеттеседі және ядродағы Nrf2 белсенділігін арнайы тежейді. Сонымен қатар, пероксисома пролифераторымен белсендірілген рецептор-?, ER?, эстрогенмен байланысты рецептор-? және глюкокортикоидты рецепторлар сияқты басқа ядролық рецепторлар да Nrf2 белсенділігінің эндогендік тежегіштері болып табылады [9], [10].
Nrf2/HO-1 осінің қабынуға қарсы рөлі
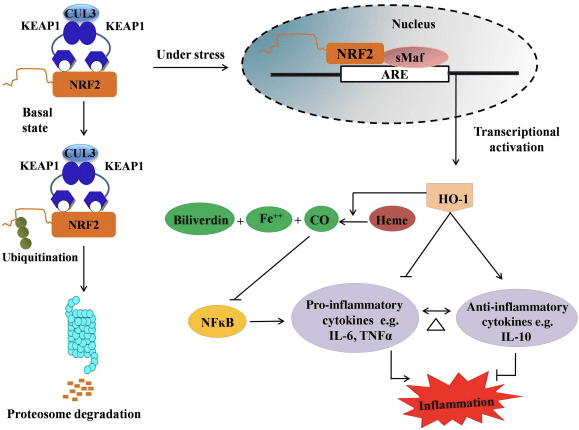
HO-1 – гемнің көміртегі тотығы (СО) мен бос темірге, ал биливердиннің билирубинге дейін ыдырауын катализдейтін индукцияланатын изоформа және жылдамдықты шектейтін фермент. Қабынуға қарсы бос гемнің ферментативті ыдырауы, сонымен қатар СО және билирубин сияқты қабынуға қарсы қосылыстардың өндірісі HO-1 қорғаныс әсерін сақтауда маңызды рөл атқарады (2-сурет).
2-сурет Nrf2/HO-1 жолына шолу. Базальды жағдайларда Nrf2 өзінің Keap1 репрессорымен байланысады, бұл убиквитинацияға, содан кейін протеазоманың деградациясына әкеледі. Тотығу стрессі кезінде бос Nrf2 ядроға ауысады, онда ол шағын Maf отбасы мүшелерімен димеризацияланады және HO-1 сияқты ARE гендерімен байланысады. Жоғары реттелген HO-1 гемді СО, билирубин және бос темірге катализдейді. CO NF-?B жолының ингибиторы ретінде әрекет етеді, бұл қабынуға қарсы цитокиндердің экспрессиясының төмендеуіне әкеледі, ал билирубин антиоксидант ретінде де әрекет етеді. Сонымен қатар, HO-1 қабынуға қарсы цитокиндерді тікелей тежейді, сонымен қатар қабынуға қарсы цитокиндерді белсендіреді, осылайша қабыну процесінің теңгеріміне әкеледі.
Nrf2 mRNA және ақуыз экспрессиясын арттыру арқылы HO-1 генін индукциялайды және ол көптеген in vitro және in vivo зерттеулерінде кеңінен қолданылатын классикалық Nrf2 реттелетін гендердің бірі болып табылады. Бірнеше зерттеулер HO-1 және оның метаболиттерінің Nrf2 арқылы болатын қабынуға қарсы маңызды әсерлері бар екенін көрсетті. Белсендірілген Nrf1 арқылы жүзеге асырылатын HO-2 экспрессиясының жоғарылауы NF?B сигналының тежелуіне әкеледі, нәтижесінде ішек шырышты қабатының зақымдануы және ерлердің Спраге-Доули егеуқұйрықтарының бауыр трансплантациясы моделінде тығыз байланыс дисфункциясы төмендейді [35]. Nrf2-тәуелді HO-1 экспрессиясының жоғарылауы тінтуірдің C2C12 миобласттарын H2O2 цитотоксикалық әсерінен қорғай алады [36]. Nrf2-тәуелді HO-1 RAW264.7- немесе тінтуірдің перитонеальді макрофагынан алынған көбік жасушаларының макрофагтарында липополисахарид (LPS) арқылы болатын қабыну реакцияларына әсер етеді. Nrf2 белсенділігі көбік жасушаларының макрофагтарының фенотипін десенсибилизациялайды және атеросклероздың өршуінде маңызды рөл атқаратын макрофагтардың шамадан тыс қабынуын болдырмайды [37]. Nrf2/HO-1 осі LPS индукцияланған тінтуірдің BV2 микроглиальды жасушаларына және тінтуірдің гиппокампалық HT22 жасушаларына әсер етеді, бұл нейроинфламацияға әсер етеді. Тінтуірдің гиппокампалы HT1 жасушаларының жасуша өлімін қорғайтын тінтуірдің BV2 микроглиальды жасушаларында Nrf2 жолы арқылы HO-22 экспрессиясының жоғарылауы [38]. Сонымен қатар, Nrf2 индукторын көміртегі тотығы (CO) шығарғышымен біріктіретін кобальт негізіндегі гибридті молекулалар (HYCO) Nrf2/HO-1 экспрессиясын арттырады, СО-ны босатады және in vitro қабынуға қарсы белсенділік көрсетеді. Сондай-ақ HYCOs тіндердің HO-1 деңгейін жоғарылатады және in vivo енгізгеннен кейін қанға CO39 жеткізеді, бұл олардың қабыну жағдайларына қарсы әлеуетті қолданылуын қолдайды [2]. Nrf1/HO-40 жоғарылауы таурин хлораминдерімен өңделген тышқан макрофагтарының эффероцитарлық белсенділігін арттыру арқылы қабынуды азайтады [2]. Барлығы жоғарыда түсіндірілген эксперименттік модельдер Nrf1/HO-2 осінің қабынуға қарсы функцияда үлкен рөл атқаратынын көрсетті, бұл NrfXNUMX қабынумен байланысты ауруларда терапевтік мақсат екенін көрсетеді.
Сонымен қатар, CO, билирубин сияқты HO-1 жанама өнімдері тотығу стрессі және жасушалардың зақымдалуы кезінде күшті антиоксидант ретінде әрекет етеді [41], [42]; ол аутоиммунды энцефаломиелит пен гепатитті басады [43], [44]; және iNOS және NO генерациясының алдын алу арқылы тышқандар мен егеуқұйрықтарды эндотоксикалық шоктан қорғайды [45], [46], [47]. Сонымен қатар, билирубин эндотелийдің белсендіруін және дисфункциясын төмендетеді [48]. Бір қызығы, билирубин адгезия молекуласы-1 арқылы эндотелий лейкоциттерінің трансмиграциясын азайтады [49]. HO-1 күшті қабынуға қарсы агент ретінде ғана емес, сонымен қатар оның метаболиттерін де көрсететін бұл арнайы сілтемелер.
Қабыну медиаторлары және Nrf2 тежейтін ферменттер
Цитокиндер және хемокиндер
Цитокиндер әртүрлі жасушалардан бөлінетін төмен молекулалық ақуыздар мен полипептидтер; олар жасушаның өсуін, дифференциациясын және иммундық функцияны реттейді және қабыну мен жараларды емдеуге қатысады. Цитокиндерге интерлейкиндер (ИЛ), интерферондар, ісік некрозының факторы (ТНФ), колонияны ынталандырушы фактор, хемокиндер және өсу факторлары жатады. Кейбір цитокиндер қабынуға қарсы медиаторлар ретінде есептеледі, ал басқалары қабынуға қарсы функцияларға ие. Тотығу стрессінің әсері мақсатты жасушаларда тотығу стрессін тудыратын цитокиндердің шамадан тыс өндірілуіне әкеледі. NF-?B тотығу стресімен белсендірілгенде бірнеше қабынуға қарсы цитокиндер артық өндіріледі. Сонымен қатар, қабынуға қарсы тотығу стресі NF-?B одан әрі белсендіруін және цитокиндердің артық өндірілуін тудырады. Nrf2/ARE жүйесін белсендіру бұл циклды бұзуда маңызды рөл атқарады. Хемокиндер - бұл қабыну жасушаларының миграциясын басқаратын негізгі рөлі шағын цитокиндердің отбасы. Олар негізінен лейкоциттер, моноциттер, нейтрофилдер және басқа эффекторлық жасушалар үшін химиатрактант ретінде қызмет етеді.
Nrf2 белсендіруі IL-6 және IL-1 қоса алғанда, қабынуға қарсы цитокиндердің LPS-индукцияланған транскрипциялық жоғарылауын болдырмайды деп хабарлады [50]. ИЛ-1? және ИЛ-6 өндірісі де Nrf2?/? декстран сульфатынан туындаған колиті бар тышқандар [51], [52]. Nrf2 төменгі ағындағы IL-17 және басқа қабыну факторлары Th1 және Th17 өндірісін тежейді және көптеген склероздың, аутоиммунды энцефалиттің тәжірибелік үлгісінде ауру процесін басады [53]. Nrf2-тәуелді антиоксидантты гендер HO-1, NQO-1, Gclc және Gclm блоктайды TNF-?, IL-6, моноциттік химиатрекциялық ақуыз-1 (MCP1), макрофагтардың қабыну протеині-2 (MIP2) және қабыну медиаторлар. Бірақ Nrf2-нокаут тышқандары жағдайында қабынуға қарсы әсер болмайды [54]. LPS-пен өңделген Nrf2-нокаут тышқандарының перитонеальді нейтрофилдерінде жабайы типті (WT) жасушаларға қарағанда цитокиндердің (TNF-? және IL-6) және химокиндердің (MCP1 және MIP2) айтарлықтай жоғары деңгейлері бар [54]. In vitro, Nrf2 генін адам мен қоянның қолқаның тегіс бұлшықет жасушаларына тасымалдау MCP1 секрециясын басады [8], [55] және Nrf2-ге тәуелді HO-1 экспрессиясы TNF-?-стимуляцияланған NF-?B және MCP-1-ді басады. адамның кіндік венасының эндотелий жасушаларында секреция [56]. Бұл нәтижелер қабыну тітіркендіргіштеріне жауап ретінде Nrf2 сигналының жоғарылауы қабынуға қарсы цитокиндер мен хемокиндердің шамадан тыс өндірілуін тежейтінін, сондай-ақ NF-?B белсендіруін шектейтінін көрсетеді.
Жасуша адгезиясының молекулалары
Жасуша адгезиясының молекулалары (CAMs) - бұл жасушалармен немесе жасушадан тыс матрицамен байланысатын ақуыздар. Жасуша бетінде орналасып, олар жасушаны тану, жасушаны белсендіру, сигнал беру, пролиферация және дифференциацияға қатысады. CAM арасында ICAM-1 және VCAM-1 иммуноглобулиндердің супертүрінің маңызды мүшелері болып табылады. ICAM-1 лейкоциттер мен эндотелий жасушаларының мембраналарында төмен концентрацияда болады. Цитокинді ынталандыру кезінде концентрация айтарлықтай артады. ICAM-1 IL-1 және TNF арқылы индукциялануы мүмкін және тамыр эндотелийімен, макрофагтармен және лимфоциттермен экспрессияланады. Бұл интегриннің лиганды, лейкоциттерде кездесетін рецептор. ICAM-1-интегрин көпірі белсендірілгенде, лейкоциттер эндотелий жасушаларымен байланысады, содан кейін субэндотелий тіндерге ауысады [57]. VCAM-1 лимфоциттердің, моноциттердің, эозинофилдердің және базофилдердің тамырлы эндотелийге адгезиясын қамтамасыз етеді және лейкоциттердің жиналуына ықпал етеді, бұл ақыр соңында тотығу стрессінен тіндердің зақымдалуына әкеледі. Nrf2 VCAM-1 промоторлық белсенділігін тежейді [58]. Nrf2 реттелетін HO-1 ағыны гені E-selektin және VCAM-1 экспрессиясына, эндотелий жасушаларымен байланысты адгезия молекулаларына әсер етуі мүмкін [59]. CD-14, TREM1, SELE, SELP және VCAM-1 сияқты бірнеше CAM-ның өкпе экспрессиясы Nrf2?/? тышқандар Nrf2+/+ тышқандарына қарағанда [60]. Адамның аорта эндотелий жасушаларындағы Nrf2 TNF-?-индукцияланған VCAM-1 экспрессиясын басады және TNF-?-индукцияланған моноциттік U937 жасушаларының адгезиясына кедергі жасайды [8]. Nrf2 шамадан тыс экспрессиясы адамның микротамырлы эндотелий жасушаларында TNF-?-индукцияланған VCAM-1 генінің экспрессиясын тежейді [61]. Табиғи антиоксидант 3-гидроксиантранил қышқылы (HA), қабыну немесе инфекция кезінде кинуренин жолы ретінде белгілі метаболикалық жолмен in vivo түзілген l-триптофан метаболиттерінің бірі, HO-1 экспрессиясын индукциялайды және адамның кіндігінде Nrf2 ынталандырады. вена эндотелий жасушалары (HUVEC). HA арқылы индукцияланған Nrf2-тәуелді HO-1 экспрессиясы MCP-1 секрециясын, VCAM-1 экспрессиясын және атеросклероздағы тамырлардың зақымдалуымен және қабынуымен байланысты NF-kB белсендіруін тежейді [56]. Пролиферативті және қабынуға қарсы синтетикалық халькон туындысы 2?,4?,6?-трис (метоксиметокси) хальконы ICAM-1, қабынуға қарсы цитокин IL-1? және TNF-? тринитробензол сульфон қышқылымен өңделген тышқандардың тоқ ішек тініндегі экспрессиясы [62]. Nrf2 жоғарылауы ликопенмен өңделген адамның ретинальды пигментті эпителий жасушаларында TNF-?-индукцияланған ICAM-1 экспрессиясын тежейді [63]. Барлық осы зерттеулер Nrf2 қабыну жасушаларының қабынған тінге көшуін және инфильтрациясын реттеу арқылы қабыну процесінде шешуші рөл атқарады деп болжайды.
Матрицалық металлопротеиназалар (ММП)
MMPs жасушадан тыс матрицада кеңінен кездеседі және жасуша пролиферациясы, миграциясы, дифференциациясы, жараларды емдеу, ангиогенез, апоптоз және ісік метастазасы сияқты физиологиялық және патологиялық процестерге қатысады. Nrf2/HO-1 осінің макрофагтардағы MMP-9-ды және адамның ішек эпителий жасушаларындағы MMP-7-ні тежейтіні хабарланды және бұл ішектің қабыну ауруларын емдеуде пайдалы [62], [64]. Ультракүлгін сәулеленуден туындаған терінің зақымдануы WT тышқандарына қарағанда Nrf2-нокаутында ауыррақ және MMP-9 деңгейі айтарлықтай жоғары, бұл Nrf2 MMP-9 экспрессиясын төмендететінін көрсетеді. Сондықтан Nrf2 ультракүлгін сәулеленуден қорғайтын құрал болып саналады [65]. Басқа зерттеу сонымен қатар ісік жасушаларының инвазиясында және қабынуында MMP-9 төмендетілген транскрипциялық белсендіру NF-kB сигналдық жолын тежеу арқылы реттелетінін хабарлады [66]. Жұлынның жарақаттануында NF-kB сигналдық жолы MMP-9 мРНҚ деңгейлерін реттеуге де қатысады [67]. Демек, қабынуда ММП реттелуіне тікелей Nrf2 жолы немесе жанама түрде Nrf2 әсер ететін NF-?B жолы әсер етеді.
Циклооксигеназа-2 (COX2) және индукцияланатын азот оксиді синтазасы (INOS)
Nrf2-нокаут тышқандарына жүргізілген эксперименттер сериясы оның қабынудағы және COX-2 және iNOS сияқты қабынуға қарсы гендердің реттелуіндегі шешуші рөлін көрсетті. Алғаш рет Хор және т.б. Nrf2?/? тоқ ішек тіндерінде COX-2 және iNOS сияқты қабынуға қарсы цитокиндердің экспрессиясының жоғарылауы туралы хабарлады? тышқандар WT Nrf2+/+ тышқандарымен салыстырылды, бұл Nrf2 олардың белсенділігін басатынын көрсетеді [51]. Айқышгүлді көкөністерде кездесетін белгілі Nrf2 активаторларының бірі болып табылатын сульфорафанмен алдын ала өңдеу туралы басқа есеп оның TNF-?, IL-1?, COX-2 және iNOS экспрессиясын иРНҚ-да тежейтін қабынуға қарсы әсерін көрсетті. және Nrf2+/+ тышқандарының бастапқы перитонеальді макрофагтарындағы ақуыз деңгейі Nrf2?/? тышқандар [68]. Сол сияқты, LPS-индукцияланған қабынуы бар Nrf2-нокаут тышқандарының гиппокампусы да iNOS, IL-6 және TNF- сияқты қабыну маркерлерінің жоғары көрінісін көрсетеді? WT тышқандарына қарағанда [69]. Сол сияқты, Nrf2-нокаут тышқандары 1-метил-4-фенил-1,2,3,6-тетрагидропиридин тудыратын тотығу стрессіне аса сезімтал, сонымен қатар COX-2, iNOS сияқты қабыну маркерлерінің мРНҚ мен ақуыз деңгейлерінің жоғарылауын көрсетеді. , IL-6 және TNF-? [70]. Сонымен қатар, Nrf2?/-ден алынған бауырлар? метионин және холин тапшылығы бар диетаға қарсы тышқандарда бір диетадағы WT тышқандарына қарағанда, Cox5 және iNOS mRNA экспрессиясы ~ 2 есе жоғары, бұл Nrf2 қабынуға қарсы рөлін көрсетеді [71]. Жақында Ким және т.б. фитохимиялық этил пируватының BV2 жасушаларында Nrf2 сигналы арқылы iNOS экспрессиясын төмендету арқылы қабынуға қарсы және тотығуға қарсы әсерлерін көрсететінін көрсетті. Олар этил пируватының Nrf2 ядролық транслокациясын индукциялайтынын көрсетті, бұл ақыр соңында p65 және p300 арасындағы өзара әрекеттесуді тежейді, бұл iNOS экспрессиясының төмендеуіне әкеледі [72]. Сонымен қатар, карбазол аналогы LCY-2-CHO Nrf2 белсендіреді және оның ядролық транслокациясын тудырады, бұл егеуқұйрық аорта тамырларының тегіс бұлшықет жасушаларында COX2 және iNOS экспрессиясының [73] басылуына әкеледі.
NLRP2 iIflammasome-белсенділігін реттеудегі Nrf3 парадоксалды рөлі
NLR отбасы, 3 (NLRP3) қабынуы бар пирин домені - бұл патогенді тану рецепторы (PRR) ретінде жұмыс істейтін және патогенмен байланысты молекулалық үлгілер (PAMPs), зақымдану сияқты микробтық, тотығу стресс сигналдарының кең ауқымын танитын көп ақуызды кешен. байланысты молекулалық үлгі молекулалары (DAMPs) және ROS [74]. Белсендірілген NLRP3 қабынуы каспаза-1 бөлінуіне және қабынуға қарсы цитокин интерлейкин-1 секрециясына делдал болады? (IL-1?) бұл, сайып келгенде, пироптоз деп аталатын жасуша өлімінің процесін тудырады, ол хосттарды патогендердің кең ауқымынан қорғайды [75]. Дегенмен, қабынудың аберрантты активтенуі трансмиссивті губка тәрізді энцефалопатиялар, Альцгеймер ауруы, Паркинсон ауруы, сондай-ақ 2 типті қант диабеті [76], қатерлі ісік [77], подагра және атеросклероз [78] сияқты ақуыздың қате қатпарлануымен байланысты.
Rong Hu тобының Nrf2-нің қабынудың теріс реттелуімен байланысы туралы жақында жүргізген бақылауы, Nrf2 NLRP1 қабынуының активтенуінің, каспазаның-3 бөлінуінің және IL-1? макрофагтарда генерациялау. Сонымен қатар, белгілі Nrf1 активаторы, терт-бутилгидрохинон (tBHQ) ARE-ны Nrf2-тәуелді жолмен белсендіру арқылы NLRP3 транскрипциясын теріс реттеді [2]. Жоғарыда аталған байқаудан басқа, сол топ диметил фумараты (DMF) Nrf79 ядролық транслокациясына және NLRP2 қабыну түйінін тежеуге қатысатын Nrf2 сигналдық жолын белсендіру арқылы DSS-индукцияланған колиттің алдын алатыны анықталды [3].
Табиғи және синтетикалық қосылыстарды қолданатын бірқатар эксперименттер сонымен қатар Nrf2-нің NLRP3 қабынуының белсендірілуіне ингибиторлық әсерін анықтады. Мысалы, қызыл жегі бар тышқандардағы эпигаллокатехин-3-галлатты (EGCG) емдеу Nrf3 сигналдық жолымен жүретін бүйрек NLRP2 қабынуының белсендірілуін төмендететінін көрсетті [81]. Сол сияқты, цитраль (3,7-диметил-2,6-октадиенал), қытайлық Litsea cubeba шөп дәрі-дәрмекіндегі негізгі белсенді қосылыс, жеделдетілген және ауыр қызыл жегі (ASLN) тінтуір үлгісіндегі Nrf3 антиоксиданттық сигнал беру жолы арқылы NLRP2 қабынуының активтенуін тежейді. [82]. Сол сияқты, биоханин Nrf2 жолын белсендіру және еркек BALB/c тышқандарында NLRP3 қабынуының белсендірілуін тежеу арқылы LPS/GalN-индукцияланған бауыр жарақатынан қорғады [83]. Сонымен қатар, мангиферин дозаға тәуелді түрде Nrf2 және HO-1 экспрессиясын жоғары реттейтіні және LPS/D-GalN индукцияланған бауыр NLRP3, ASC, каспаза-1, IL-1? және TNF-? өрнек [84].
Nrf3 арқылы NLRP2 теріс реттелуіне қарамастан, ол NLRP3 және AIM2 қабыну функциясын да белсендіреді. Хайтао Вэн және оның әріптестері Nrf2 ?/? тінтуірдің макрофагтары NLRP3 және AIM2 қабынуының ақаулы активтенуін көрсетті, бірақ NLRC4 қабынуы емес [85]. Бір қызығы, бұл бақылау Nrf2-нің қабынумен байланысты аурулар контекстіндегі белгісіз функцияларын бейнелейді; сондықтан оны терапевтік мақсат ретінде қарастырмас бұрын Nrf2 қабыну функциясын белсендіретін механизмді ашу үшін әрі қарай зерттеу өте маңызды.
Nrf2 арқылы қабынуға қарсы цитокин транскрипциясын басу
Хроматиндік иммунопреципитация (ChIP)-seq және ChIP-qPCR нәтижелеріне негізделген өте жақында жүргізілген зерттеу тінтуірдің макрофагтарына әкелді. және РНҚ Пол II тартуды тежейді. Нәтижесінде РНҚ Пол II ИЛ-2 және ИЛ-6 транскрипциялық активтенуін өңдей алмайды? бұл ген экспрессиясының тежелуіне әкеледі. Масаюки Ямамото тобы алғаш рет Nrf1 өзінің төменгі гендерін ARE арқылы транзакциялап қана қоймай, сонымен қатар РНҚ Pol II тартылуын тежеу арқылы ARE бар немесе онсыз нақты гендердің транскрипциялық активтенуін басатын жаңа механизмді ашты [6].
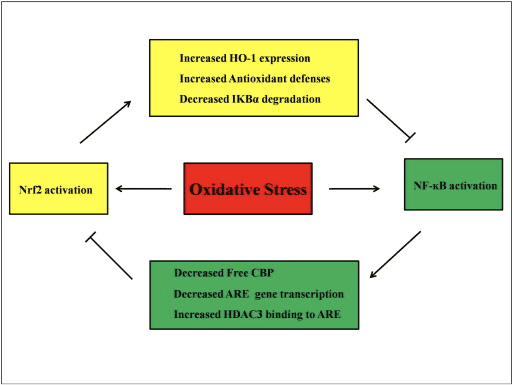
Nrf2 және NF-?B жолдары арасындағы өзара байланыс
NF-?B - жануарлар жасушаларының барлық дерлік түрлерінде кездесетін және қабыну, апоптоз, иммундық жауап, жасушалардың өсуі және дамуы сияқты әртүрлі процестерге қатысатын ДНҚ транскрипциясына жауапты ақуыз кешені. p65, NF-?B отбасының Rel протеині, трансактивация доменіне ие, ал p50 транскрипцияны белсендіру үшін Rel ақуызымен гетеродимеризацияны қажет етпейді және қажет етеді. Тотығу стрессі кезінде I?B киназа (ИКК) белсендіріледі және I?B фосфорлануын тудырады, нәтижесінде NF-?B-ның бөлінуі және ядролық транслокациясы жүреді. NF-?B IL-6, TNF-?, iNOS, IL-1 және жасушаішілік адгезиясы COX-2 сияқты қабынуға қарсы медиаторлардың транскрипциясын тудырады.
NF-?B нормадан тыс реттелуі ревматоидты артритпен, астмамен, ішектің қабыну ауруымен және Helicobacter pylori инфекциясынан туындаған гастритпен байланысты болды [86]. Қазіргі уақытта NF-kB белсенділігі Keapl/Nrf2/ARE сигналдық жолына негізінен үш аспектіде әсер етеді деп саналады: біріншіден, Keap1 IKK-ны төмендетеді? ubiquitination арқылы, осылайша NF-?B белсенділігін тежейді [87]. Екіншіден, қабыну процесі циклопентенон простагландин 2d-PGJ15-ден алынған COX2 сияқты қабыну медиаторларын, Keap1-пен әрекеттесетін және Nrf2-ні белсендіретін күшті электрофилді тудырады, осылайша NF-kB белсенділігінің бір мезгілде тежелуімен ген транскрипциясын бастайды [58], [88] 3 A, B суреті). Үшіншіден, NF-?B бәсекеге қабілетті Nrf2 транскрипциялық коактиваторымен CBP [89], [90] біріктіре алады (Cурет 3 C, D).
3-сурет Nrf2 және NF-?B жолдары арасындағы қиылысу. (A) Keap1 IKK-ны CUL3-делдалдық убиквитинациясына және протеазоманың деградациясына бағыттайды, бұл ақыр соңында NF-?B фосфорлануының тежелуіне әкеледі және бұл механизм сонымен қатар Nrf2 және IKK-ның Keap1-мен бәсекелес байланысуы ретінде жұмыс істейді. (B) Тотығу стрессі NF-?B фосфорланатын IKK-ны белсендіреді, бұл оның ядроға транслокациясына және COX-2 сияқты қабынуға қарсы цитокиндердің белсендірілуіне әкеледі. 2d-PGJ15 деп аталатын COX-2 терминалдық өнімі Nrf2 индукторы ретінде әрекет етеді, ол ақырында тотығу стрессін басуға әкеледі. (C) Nrf2 ARE басқаратын ген экспрессиясын бастау үшін шағын Maf және басқа транскрипциялық аппараттармен бірге CBP транскрипциялық кофакторымен байланысады. (D) NF-?B CBP-мен бәсекеге қабілетті түрде байланысқанда, ол CBP-нің Nrf2-мен байланысуын тежейді, бұл Nrf2 трансактивациясының тежелуіне әкеледі.
Nrf2 және NF-?B сигналдық жолдары төменгі ағындағы мақсатты ақуыздардың транскрипциясын немесе қызметін басқару үшін өзара әрекеттеседі деп болжанады. Бұл болжамды негіздеу үшін көптеген мысалдар Nrf2 және NF-?B жолдарының мүшелері арасында тікелей немесе жанама белсендіру мен тежелу орын алатынын көрсетеді (Cурет XNUMX). 4). LPS-ке жауап ретінде Nrf2 нокдаун NF-?B транскрипциялық белсенділігін және NF-?B-тәуелді ген транскрипциясын айтарлықтай арттырады, бұл Nrf2 NF-?B белсенділігіне кедергі келтіретінін көрсетеді [60], [91]. Сонымен қатар, Nrf2-тәуелді төмен HO-1 экспрессиясының жоғарылауы NF-?B белсенділігін тежейді. Қуық асты безінің қатерлі ісігінің жасушалары ?-тохоферилсукцинатқа, Е витаминінің туындысына қысқа әсер еткенде, HO-1 экспрессиясы жоғарылайды. HO-1 соңғы өнімдері NF-?B ядролық транслокациясын тежейді [92]. Бұл in vivo зерттеулер Nrf2 NF-kB сигналдық жолын теріс реттейтінін көрсетеді. LPS NF-?B ДНҚ байланыстыру белсенділігін ынталандырады және NF-?B-ның p65 суббірлігінің деңгейі Nrf2?/? өкпеден алынған ядролық сығындыларда айтарлықтай жоғары. WT тышқандарына қарағанда, NF-?B активациясында Nrf2 теріс рөлін көрсетеді. Оның үстіне Nrf2?/? LPS және TNF-мен өңделген тышқан эмбрионының фибробласттары? IKK активациясынан туындаған NF-?B белсенділенуін және I?B-? деградация [60]. Ал респираторлық синцитиальды вирус клиренсі айтарлықтай төмендейді, ал NF-?B ДНҚ-байланыстыру белсенділігі Nrf2?/? тышқандар WT тышқандарымен салыстырғанда [93]. Nrf2?/-да пристаннан туындаған қызыл жегі нефриті? сульфорафанмен бірге емделген тышқандарда бүйректің ауыр зақымдануы және патологиялық өзгерістері, сондай-ақ WT-мен салыстырғанда iNOS экспрессиясының жоғарылауы және NF-?B белсендіруі бар, бұл Nrf2 NF-?B сигналдық жолын тежеу және ROS тазарту арқылы қызыл жегінің нефритін жақсартады деп болжайды [94] ]. NF-?B белсенділігі жасушаларды LPS және TNF-?-мен бірге Nrf2 индукторымен өңдегенде де пайда болады. Мысалы, синтетикалық халькон туындысы адам ішек эпителийінің HT-1 жасушаларында HO-29 экспрессиясын индукциялау арқылы TNF-?-индукцияланған NF-?B белсендіруін тікелей және жанама және ішінара тежейді [62]. F344 егеуқұйрықтарын 3H-1,2-дитиол-3-тионмен (D3T) өңдеген кезде NF-?B транслокациясының және ДНҚ-байланыстыру белсенділігінің басылуы, сондай-ақ гепатоциттерде iNOS экспрессиясының басылуы анықталады [95]. Сульфорафанмен және LPS-пен бірге өңдеуден кейін LPS-индукцияланған iNOS, COX-2 және TNF-? шикі 264.7 макрофагтардың реттелуі төмендейді, сульфорафанның NF-?B ДНҚ байланысуын тежеу арқылы қабынуға қарсы белсенділігі бар деп болжайды [96]. Nrf2 және NF-?B жолдары арасындағы байланысты түсіндіру үшін бірнеше эксперименттік зерттеулер жүргізілгенімен, қарама-қайшы нәтижелер сақталады. Nrf2 және NF-kB [97] арасында оң және теріс реттеулер туралы хабарланды. Әдетте, химиопревентивтік электрофилдер 3H-1,2-дитиол-3-тион, сульфорафан және тритерпеноидты CDDO-Me NF-kB және оның төмендетілген гендерін тежеу арқылы Nrf2 белсендіреді [98], [99], [100]. Керісінше, ROS, LPS, ағынның ығысу кернеуі, тотыққан LDL және темекі түтіні сияқты бірнеше агенттер немесе жағдайлар Nrf2 және NF-kB белсенділігін арттырады [97]. Сонымен қатар, in vivo зерттеулер Nrf2?/?-ден бөлінген бауырда NF-kB белсенділігінің төмендейтінін анықтады. тышқандар мен NF-?B байланысу белсенділігі Nrf2?/? Nrf2+/+ тышқандарына қарағанда [101]. Дегенмен, Nrf2 аденовирустық векторымен өңделген адамның аорта эндотелий жасушалары NF-?B белсенділігіне әсер етпей, NF-?B ағынының гендерін тежейді [8].
4-сурет Nrf2 және NF-?B реттеуші циклі. Nrf2 жолы I?B-? деградациясының алдын алу арқылы NF-?B активациясын тежейді. және HO-1 экспрессиясын және ROS және детоксикациялайтын химиялық заттарды бейтараптандыратын антиоксиданттық қорғанысты арттыру. Нәтижесінде, ROS-пен байланысты NF-?B белсендіру басылады. Сол сияқты, NF-?B-делдалдық транскрипция Nrf2 активациясын азайту арқылы азайтады ARECBP үшін Nrf2-мен бәсекелесе отырып, геннің транскрипциясы және бос CREB байланыстыратын ақуыз. Сонымен қатар, NF-?B ARE аймағына гистон-деацетилазаның (HDAC3) тартылуын арттырады және осылайша Nrf2 транскрипциялық активациясының алдын алады.
Nrf2 сигналдық жолын белсендіру адам ағзасындағы жасушалардың антиоксиданттық қабілетін арттыру арқылы реактивті тотықтырғыштарды детоксикациялауға қатысатын ферменттер мен гендердің экспрессиясында үлкен рөл атқарады. Бүгінгі таңда көптеген зерттеу зерттеулері бар болғанымен, Nrf2 белсендірудегі реттеу механизмдері толық түсінілмеген. Nrf2 сигналдық жолының қабынуды емдеудегі ықтимал рөлі де табылды. Доктор Алекс Хименес DC, CCST Insight
Қабыну ауруларындағы Nrf2 рөлі
In vivo зерттеулер Nrf2 әртүрлі жүйелерге әсер ететін қабыну ауруларында маңызды рөл атқаратынын көрсетті; оларға гастрит, колит, артрит, пневмония, бауырдың зақымдануы, жүрек-қан тамырлары аурулары, нейродегенеративті ауру және мидың зақымдалуы жатады. Бұл зерттеулерде Nrf2?/? жануарлар WT жануарларына қарағанда қабынудың және тіндердің зақымдалуының ауыр белгілерін көрсетті. Сондықтан Nrf2 сигналдық жолы қабыну ауруларында қорғаныс әсері бар деп саналады. Шошқаның панкреатикалық эластазасының трахея ішілік орнатылуы өкпенің созылмалы обструктивті ауруын, әсіресе эмфиземаны тудырады. Nrf2 жетіспейтін тышқандар эмфиземаға өте сезімтал және альвеолярлы макрофагтарда HO-1, PrxI және антипротеаза генінің SLPI экспрессиясының төмендеуі байқалады. Nrf2 өкпенің зақымдалуына қарсы макрофагтық қорғаныс жүйесіндегі негізгі реттеуші болып саналады [102]. 2 ай бойы темекі түтінінің әсерінен туындаған эмфиземасы бар Nrf6 тапшылығы бар тышқандар бронх-альвеолярлық қабынудың жоғарылауын, альвеолалардағы тотығу стресс маркерлерінің жоғарылауын және альвеолярлы септал жасушаларының апоптозының жоғарылауын көрсетеді, бұл Nrf2 темекі тудырған антиоксидант экспрессиясының жоғарылауы арқылы әсер етеді деп болжайды. гендер [102], [103]. Nrf2 бұзылған кезде, аллергенмен байланысты тыныс жолдарының қабынуы және сопақ бумин кешенін пайдаланатын астма тыныс алу жолдарының қабынуын, тыныс жолдарының гиперреактивтілігін, бокал жасушаларының гиперплазиясын және бронх-альвеолярлы жуу мен спленоциттерде Th2 жоғары деңгейлерін көрсетеді, ал Nrf2-делдалдық сигнал беру жолын шектейді. , шырыштың гиперсекрециясы және тыныс жолдарының гиперреактивтілігі, сонымен қатар астманың дамуына жол бермейтін көптеген антиоксиданттық гендерді индукциялау [104]. Плевра қуысына каррагенан инъекциясы плеврит тудырады, ал Nrf15 қабыну жасушаларында 2d-PGJ2 жинақталуы тінтуірдің перитонеальді макрофагтарымен шектеледі. Қабынудың бастапқы кезеңінде 15d-PGJ2 Nrf2 белсендіреді және HO-1 және PrxI индукциясы арқылы қабыну процесін реттейді. Зерттеу сонымен қатар COX-2 15d-PGJ2 [105] өндірісі арқылы ерте фазада қабынуға қарсы әсерге ие екенін көрсетті. Натрийдің 1% декстран сульфатын 1 апта бойы ішке қабылдау крипттердің қысқаруын және тоқ ішек тініндегі қабыну жасушаларының инфильтрациясын қамтитын гистологиялық өзгерістермен байланысты колитті индукциялайды. Колит кезінде ішектің тұтастығын қорғау үшін Nrf2 қабынуға қарсы цитокиндерді реттеу және II фазаны детоксикациялаушы ферменттерді индукциялау арқылы маңызды рөл атқара алады [51]. LPS-индукцияланған өкпе сепсисінің Nrf2-нокауттық тінтуір үлгісінде NF-?B белсенділігі COX-2, IL-113, IL-6 және TNF сияқты қабыну цитокиндерінің әсерін реттейді? қабынуды бастау және ілгерілету үшін маңызды [60]. Nrf2 осы қабыну факторларын реттеу арқылы қабыну зақымдануын азайтады. Жедел қабынудың осы үлгілерінде антиоксиданттық ферменттердің, қабынуға қарсы цитокиндердің және медиаторлардың Nrf2 сигналдық жолы арқылы реттелуінің жоғарылауы WT жануарларындағы қабыну жарақатын азайтады. Бір қызығы, бұл WT тышқандарымен салыстырғанда симптомдары айтарлықтай күшейетін Nrf2-нокаут тышқандарында да хабарланды.
Nrf2-тәуелді қабынуға қарсы препараттарды зерттеу
Қорытындылай келе, біз Nrf2 сигнал жолы қабынудың көптеген салаларында реттеуші рөл атқаратынын көрсететін эксперименттерді талқыладық, сондықтан Nrf2-ге тәуелді қабынуға қарсы агенттер қабыну ауруларын емдеу үшін маңызды.
Өсімдіктер Nrf2 транскрипция факторын белсендіретін, цитопротекторлық гендердің жоғары реттелуіне әкелетін қосылыстардың ерекше бай көздері болды. Жақында әртүрлі қабынуға қарсы агенттердің, негізінен өсімдік тектес заттардың әсерін зерттеу үшін бірнеше зерттеулер жүргізілді. Мысалы, куркумин куркуманың белсенді ингредиенті болып табылады және зімбірде де аз мөлшерде кездеседі; изотиоцианаттар, атап айтқанда фенилизотиоцианаттар брокколиден, балдыркөктен және басқа да көкөністерден; ал антоцианиндер жидектер мен жүзімдер [124]. Зерттеулер көрсеткендей, бұл агенттердің барлығы жақсы антиоксиданттар ғана емес, сонымен қатар Nrf2 индукциясы арқылы күшті қабынуға қарсы әсерлері бар [125], [126]. Сондықтан өсімдік сығындысынан жаңа қабынуға қарсы Nrf2 белсендіргіштерін жасау медициналық зерттеулерде үлкен қызығушылық тудырды.
Соңғы жылдары осы қосылыстардың әрекетін растау үшін жануарларға көптеген эксперименттер жүргізілді. Артесунат негізінен ауыр безгек, церебральды безгек және ревматикалық аутоиммунды аурулар үшін қолданылады; бұл өкпенің септикалық зақымдалуында да тиімді. Артесунат Nrf2 және HO-1 экспрессиясын белсендіреді, ал соңғысы қабынудың алдын алу үшін тінге қабынуға қарсы цитокиндер мен лейкоциттердің түсуін азайтады [127]. Oryza sativa күрішінің қабығынан алынған изовитексиннің қабынуға қарсы және антиоксиданттық қасиеттері бар деп саналады; ол Nrf2/HO-1 жолын белсендіру және MAPK және NF-?B [128] тежеу арқылы LPS-индукцияланған жедел өкпе жарақатына қарсы қорғаныс рөлін атқарады. Фимасартан, ренин-ангиотензин жүйесіне әсер ететін жаңадан танымал ангиотензин II рецепторларының блокаторы қан қысымын төмендетеді; Хирургиялық индукцияланған бір жақты несепағар обструкциясы бар тышқандарды емдеу үшін фимасартанды қолдану Nrf2 және антиоксиданттық жолды жоғарылату және RAS және MAPKs [129] тежеу арқылы тотығу стрессін, қабынуды және фиброзды азайтады. Саппанон Оңтүстік-Шығыс Азияда кеңінен таралған, онда ол тұмауға қарсы, аллергияға қарсы және нейропротекторлық дәрі ретінде қолданылады; ол Nrf2 белсендіреді және NF-?B тежейді, сондықтан Nrf2- және/немесе NF-?B байланысты ауруларды емдеуде пайдалы болуы мүмкін [130]. Bixin orellana тұқымынан алынған биксин Мексика мен Оңтүстік Америкада жұқпалы және қабыну ауруларында қолданылады; ол вентиляциядан туындаған өкпе жарақатын жеңілдету және қалыпты өкпе морфологиясын қалпына келтіру үшін Nrf2-тәуелді түрде қабыну медиаторларын, альвеолярлық капиллярлардың ағуын және тотығу зақымдануын азайтады [131]. Эпигаллокатехин галлаты, сульфорафан, ресвератрол, ликопен және жасыл шай сығындысы сияқты басқа өсімдік қосылыстары Nrf2 сигналдық жолы арқылы қабыну ауруларына емдік әсер етеді [132], [133], [134]. Жақында цитрус жемістерінде болатын басқа фитохимиялық, эродидиктиолдың Nrf2 реттеу, NF-?B тежеу және тежеу арқылы цисплатин тудырған бүйрек жарақатына және сепсис тудырған жедел өкпе жарақатына қабынуға қарсы және антиоксиданттық әсерлері бар екені хабарланды. макрофагтардағы цитокиндердің экспрессиясы [135], [136]. Дегенмен, көптеген фитохимиялық заттар адамның әртүрлі ауруларының алдын алу мен емдеуге үлкен үміт береді, ал кейбіреулері клиникалық сынақ кезеңіне енді (2-кесте).
Бұл өсімдік қосылыстары Nrf2 сигналдық жолын негізінен Keap1 цистеинінің қалдықтарын өзгертетін электрофильді материалдар түрінде белсендіреді, бұл ARE-мен бос ядролық Nrf2 байланысуына әкеледі, нәтижесінде сәйкес геннің транскрипциясы белсендіріледі.
Сульфорафан және оның қатерлі ісікке, өлімге, қартаюға, ми мен мінез-құлыққа, жүрек ауруына және басқаларына әсері
Изотиоцианаттар сіздің диетаңызға түсетін ең маңызды өсімдік қосылыстарының бірі болып табылады. Бұл бейнеде мен олар үшін жасаған ең жан-жақты жағдайды жасаймын. Қысқаша назар аударыңыз ба? Төмендегі уақыт нүктелерінің бірін таңдап, сүйікті тақырыпқа өтіңіз. Толық уақыт шкаласы.
Негізгі бөлімдер:
00: 01: 14 - қатерлі ісік және өлім
00: 19: 04 - қартаю
00: 26: 30 - Ми және мінез
00: 38: 06 - Соңғы қорытынды
00: 40: 27 - Доза
Толық уақыт шкаласы:
00: 00: 34 - бейнефильмнің басты тақырыбы - сульффоран енгізу.
00: 01: 14 - Өсімдік тұтыну және барлық өлім-жітімде қысқарту.
00: 02: 12 - простата қатерлі ісігі.
00: 02: 23 - Мочевина рагы қатер.
00: 02: 34 - темекі шегудегі қатерлі ісік.
00: 02: 48 - Сүт безі қатерлігі қатері.
00: 03: 13 - Гипотетикалық: егер сізде рак бар болса? (интервенционалдық)
00: 03: 35 - қатерлі ісік пен өлім қауымдасқан деректерді басқаратын шынайы механизм.
00: 04: 38 - сульфорфан және қатерлі ісік.
00: 05: 32 - егеуқұйрықтардағы ісік ісігін дамытуға арналған брокколи өсімдік сығындысының күшті әсерін көрсететін жануарлы дәлел.
00: 26: 30 - мидың және мінез-құлық бөлімінің басы.
00: 27: 18 - аутизмге арналған брокколи өсімдік сығындысының әсері.
00: 27: 48 - глюкурофенияларға арналған глюкурофанның әсері.
00: 28: 17 - Депрессияның талқылануын бастау (ықтимал механизм мен зерттеулер).
00: 31: 21 - Стресс-индуцирленген депрессияның 10 түрлі модельдерін қолданып тышқандарды зерттеу, сульфарфетан сияқты флоксин (prozac) сияқты тиімді.
00: 32: 00 - зерттеу тышқандарда глюкарофаниннің тікелей ішуін көрсетеді, социологиялық жеңілдетудің стресс-моделінен депрессияны болдырмау үшін де тиімді.
00: 33: 01 - нейродегенерация бөлімінің басы.
00: 33: 30 - сульфорфан және Альцгеймер ауруы.
00: 33: 44 - Сульфорфан және Паркинсон ауруы.
00: 33: 51 - Сульфорфан және Хантингтон ауруы.
00: 34: 13 - Сульфорфан жылу әсерін арттырады.
00: 34: 43 - бас миының зақымдану секциясының басы.
00: 35: 01 - Туберкулезге қарсы тестілеуден кейін бірден сульфорфан препараты (тышқанның зерттеуі).
00: 35: 55 - сульфорфан және нейрондық пластика.
00: 36: 32 - Сулфорафан тышқандар типіндегі II типті қант диабеті моделін үйренуді жақсартады.
00: 37: 19 - сульфорфан және дученне бұлшықет дистрофиясы.
00: 38: 06 - Кешіктіріп бейнелеу: өлім және қатерлі ісік, ДНҚ зақымдануы, тотығу стресті және қабыну, бензолдың экскрециясы, жүрек-қан тамырлары аурулары, II типті диабет, миға әсер ету (депрессия, аутизм, шизофрения, нейрогенерация), NRF2 жолдары.
00: 40: 27 - брокколи өскіндер немесе сульффоранның дозасын анықтау туралы ойлар.
00: 41: 01 - Үйде өсіруге арналған анекдоттар.
00: 43: 14 - Пісіру температуралары және сульфарфан белсенділігі туралы.
00: 43: 45 - Глюкорафаниннен сульффоранның Gut бактерияларына айналуы.
00: 44: 24 - Көкөністерден белсенді мирозиназмен үйлескен кезде қоспалар жақсы жұмыс істейді.
00: 44: 56 - пісіру әдісі және крест тәрізді көкөністер.
Қазіргі уақытта көптеген зерттеулер Nrf2/Keap1/ARE сигналдық жолының қабынудағы рөліне бағытталған. Nrf2 арқылы реттелетін ферменттердің ішінде HO-1 стресске жауап беретін репрезентативті ферменттердің бірі болып табылады. HO-1 көрнекті қабынуға қарсы және антиоксиданттық қасиеттерге ие. Тұтастай алғанда, Nrf2 сигналдық жолы цитокиндерді, химокинді босату факторларын, ММП-ларды және басқа қабыну медиаторлары COX-2 және iNOS өндірісін теріс реттейді, олар NF-kB және MAPK тиісті жолдарына және қабынуды бақылайтын басқа желілерге тікелей немесе жанама әсер етеді. Nrf2 және NF-?B сигналдық жолдары төменгі ағындағы мақсатты ақуыздардың транскрипциясын немесе қызметін реттеу үшін өзара әрекеттеседі деп болжанады. Nrf2 арқылы NF-?B-делдалдық транскрипциялық белсенділікті басу немесе инактивациялау қабынудың ерте фазасында орын алуы мүмкін, өйткені NF-?B қабынуға қарсы медиаторлар массивінің жаңа синтезін реттейді. Дегенмен, Nrf2 мен JAK/STAT сияқты басқа сигналдық жолдар арасында байланыс бар ма, қабыну кезінде табиғи өсімдік көздерінен алынған Nrf2 активаторларының маңызы және биологиялық белсенділікті қалай жақсарту керек сияқты зерттеулерде әлі де кейбір шектеулер бар. және осы қосылыстардың мақсаттылығын күшейту. Бұлар қосымша эксперименттік тексеруді қажет етеді.
Сонымен қатар, Nrf2 сигналдық жолы > 600 генді [163] реттей алады, оның ішінде > 200 цитопротекторлық ақуыздарды [164] кодтайды, олар да қабыну, қатерлі ісік, нейродегенеративті аурулар және басқа да негізгі аурулармен [165] байланысты. Көптеген қатерлі ісік ауруларында Nrf2 сигнал беру жолы реттелмейтінін көрсететін өсуде дәлелдемелер Nrf2 тәуелді гендік батареяның аберрантты экспрессиясына әкеледі. Сонымен қатар, қабыну тотығу стрессімен байланысты ауруларда, әсіресе қатерлі ісік ауруларында маңызды рөл атқарады. Қабынуға қарсы тұру үшін бірнеше Nrf2 активаторларын қолдану онкогенезді индукциялайтын Nrf2 гендерінің аберрантты экспрессиясына әкелуі мүмкін және химиотерапияға және/немесе радиотерапияға төзімділікті тудырады. Сондықтан оның плейотропты әсерін азайту үшін Nrf2 жоғары спецификалық активаторлары жасалуы мүмкін. Nrf2 бірнеше активаторлары тотығу стрессіне байланысты ауруларда қабынуға қарсы функциялардың айтарлықтай жақсарғанын көрсетті. FDA мақұлдаған және көптеген склероз (MS) сияқты қабыну ауруларын емдеу үшін кеңінен қолданылатын Nrf2 активаторының ең жақсы мысалы - диметил фумараты. Tecfidera (Dimetylfumarate Biogen компаниясының тіркелген атауы) көптеген науқастарда склероздың қайталанатын түрлерін емдеу үшін тиімді қолданылады [152]. Дегенмен, Nrf2 активаторларын қабыну ауруларын емдеу үшін пайдалану тиімділігі Nrf2 зиянды әсерлерін болдырмау үшін қосымша тексеруді қажет етеді. Сондықтан Nrf2 арқылы қабынуға қарсы белсенділікке арналған терапияны әзірлеу маңызды клиникалық әсер етуі мүмкін. Бүкіл әлем бойынша Nrf2 сигнал беру жолының үздіксіз зерттеулері қабыну симптомдарын бақылау, қатерлі ісік, нейродегенеративті және басқа да негізгі аурулардың алдын алу және емдеу үшін жоғары мақсатты терапиялық агенттерді әзірлеуге арналған.
Қорытындылай келе, Nrf2 адам ағзасындағы тотығу стрессінің деңгейін сезінеді және сайып келгенде, антиоксидантты және детоксикациялаушы ферменттер мен гендердің реттелуіне ықпал етеді. Тотығу стресс деңгейінің жоғарылауынан туындаған созылмалы қабыну нейродегенеративті аурулармен байланысты болғандықтан, Nrf2 маңызды рөл атқара алады Альцгеймер ауруы сияқты денсаулық мәселелерін емдеуде, т.б. Біздің ақпаратымыздың көлемі хиропротикалық және омыртқаның денсаулығына қатысты мәселелермен шектеледі. Тақырыпты талқылау үшін доктор Хименеске хабарласыңыз немесе бізге телефон арқылы хабарласыңыз915-850-0900 .
Қосымша тақырыпты талқылау: Тікелей ауырсынуды хирургсыздандыру
Тізедегі ауырсыну - бұл белгілі симптом, ол әртүрлі тізе жарақаттарына және / немесе жағдайларға байланысты болуы мүмкін, соның ішіндеспорттық жарақаттар. Тізе - адам ағзасындағы ең күрделі буындардың бірі, өйткені ол төрт сүйектің, төрт байламның, әртүрлі сіңірлердің, екі менисканың және шеміршектің қиылысуынан тұрады. Американдық жанұялық дәрігерлер академиясының мәліметі бойынша, тізедегі ауырсынудың ең көп тараған себептеріне пателлярлық сублаксация, пателлярлық тендинит немесе секіргіш тізе және Осгуд-Шлаттер ауруы жатады. Тізедегі ауырсыну 60 жастан асқан адамдарда жиі кездесетініне қарамастан, тізе ауруы балалар мен жасөспірімдерде де болуы мүмкін. Тізедегі ауырсынуды RICE әдістері бойынша үйде емдеуге болады, дегенмен ауыр тізе жарақаттары шұғыл медициналық көмекті, соның ішінде хиропротикалық күтімді қажет етуі мүмкін. �
Альцгеймер ауруы және Паркинсон ауруы сияқты нейродегенеративті аурулар бүкіл әлем бойынша миллиондаған адамға әсер етеді. Бірнеше нейродегенеративті аурулардың белгілерін емдеуге арналған емдеудің әртүрлі нұсқалары бар, бірақ олардың нәтижелері жиі шектеледі. Зерттеулердің нәтижесінде ішкі және сыртқы факторлардың туындаған тотығу стрессы нейродегенеративті аурулардың дамуына себеп болуы мүмкін. The транскрипциялық фактор, Nrf2, тотығу стресінен қорғайтын негізгі қорғаныс механизмі ретінде жұмыс істеуге шешім қабылдады. Төмендегі мақаланың мақсаты - оның әсерлерін көрсету Nrf2 нейродегенеративті аурулар бойынша.
Нейродежеративті аурулар нақты ақуыздық агрегаттардың жинақталуымен байланысты, бұл жарақат алған ми мен протеостаздың жоғалуы арасындағы тығыз байланыс. Протеостаста сигналдық жолдардың, гендердің экспрессиясының және ақуызды азайту жүйелерінің реттелуін біріктіретін кең желі арқылы протеоманың молдығын және қаптауын басқаратын жасушалардың барлық процестеріне сілтеме жасайды. Бұл шолу NRF2 транскрипция коэффициентімен (ядролық фактор (эритроид туындысы 2) - ұқсас 2) қолданылатын протеостаздың трансцептуралық модуляциясы туралы ең маңызды қорытындыларды шығаруға тырысады. NRF2 классикалық түрде антиоксидантты жасушалық реакцияның негізгі реттеушісі ретінде қарастырылады, бірақ қазіргі уақытта ол протеостаздарды сақтау үшін трансекциялық машиналардың негізгі құрамдас бөлігі ретінде пайда болады. Біз NRF2-ты талқылаған кезде транскрипцияның үйлесімді және ұзаққа созылмалы реакциясын қалыптастыру үшін бұрыс ақуыздың жинақталуынан туындаған авариялық сигналдарды құрастыратын хаб ретінде қарастыруға болады. Бұл эндоплазмалық ретикулум физиологиясын, протеасомды және аутоафагиді ұстауға қатысатын гендерді бақылауға қатысты NRF2 функциясы арқылы жүзеге асырылады.
Түйінді сөздер:Нейродежеративті аурулар, Белгісіз ақуызға қарсы әрекет, Протеасом, Убицитин, Автофаги, Тоцитациялық стресс
Ядролық фактор (2) - ұқсас 2 (NRF2) - бұл клеткалық гомеостаздың негізгі реттеушісі ретінде қарастырылатын негізгі лейцинді-найзағай ақуызы. Антиоксиданттық жауап элементін (ARE) [250], [1], [2], [3], [4] деп аталатын ортақ зиянды күшейткішпен ортақ пайдаланылатын 5 гендерінің үстінен және кернеулі-индуцирленген негізін бақылайды. Бұл гендер I, II және III фазалардың детоксикациялық реакцияларына, глутатион мен пероксиреоксин / тероредоксин метаболизміне, NADPH-ны пентоздық фосфат жолымен және малик ферменттерімен, май қышқылының тотығуымен, темір метаболизмімен және протеостаздармен (6) қатысады. Бұл кең цитопротекторлық функцияларды ескере отырып, NRF2-те бір фармакологиялық шаншу созылмалы аурулардың негізгі бұзушыларының, соның ішінде тотығу, қабыну және протеотоксикалық стресстің әсерін азайтуы мүмкін. Көптеген зерттеулерде NRF2 антиоксиданттық қорғанысты және қабынуды жоюдың рөлі талқыланды (7-де қаралды). Бұл жерде протеостаздағы рөліне назар аударамыз, яғни ақуыз синтезінің, жиналуының, сатылуының және тозуының гомеостатикалық бақылауы. Мысалдар нейродегенеративті аурулар тұрғысынан қарастырылады.
Нейродегенеративті аурулардың жалпы белгісі - бұл кейбір ақуыздардың аберрантты агрегациясының пайда болуы. Осылайша, қатпарланған ақуыз агрегаттары? -Синуклеин (? -SYN) Паркинсон ауруы (ПД),? -Амилоидты (А?) Бляшкаларында және гиперфосфорланған TAU нейрофибриллярлы түйіршіктерінде Альцгеймер ауруы (AD), аңшылықта (Htt) кездеседі. Хантингтон ауруы (HD), супероксид-дисмутаза 1 (SOD1) және TAR ДНҚ байланыстыратын ақуыз 43 (TDP-43) амиотрофиялық бүйір склерозында (ALS), прион ақуызы (PrP) губка тәрізді энцефалопатияларда және т.б. Протеин агрегаттары бірнеше әсер етуі мүмкін ұялы жолдар, бұл өз кезегінде NRF2 деңгейіне және белсенділігіне әсер етуі мүмкін.
NRF2 белсенділігін реттеудің түрлі деңгейлері
Физиологиялық жағдайында жасушалар оның жылдам айналымы себебінен төмен NRF2 протеин деңгейлерін көрсетеді. Әртүрлі ынталандыруларға жауап ретінде NRF2 ақуызы жиналып, ядроға кіреді және ARE құрамындағы гендердің транскрипциясын арттырады. Сондықтан NRF2 протеинінің деңгейлерін басқару оң және теріс кіріс сигналдарын біріктіретін басты мәселе болып табылады. Бұдан әрі талқылайтын болсақ, NRF2 жылдам және тиімді жауапты ұйымдастыру үшін әртүрлі жабатын механизмдер арқылы іске қосылады, ал екінші жағынан NRF2, мүмкін, екінші фазада, оның жауапты өшіру үшін кедергі болуы мүмкін.
Классикалық тұрғыдан NRF2 активациясы тотықтырғыш немесе электрофильді қосылыстарға жасушалық реакцияның салдары ретінде қарастырылды. Осыған байланысты, убивитин E3 лигаза адаптері Келч тәрізді ECH-мен байланысқан ақуыз 1 (KEAP1) шешуші рөл атқарады. Молекулалық бөлшектер 4.1 бөлімінде қосымша қарастырылады. Қысқаша айтқанда, KEAP1 цистеиннің кристалдық қалдықтары салдарынан тотықсыздану-тотықсыздану сенсоры ретінде жұмыс істейді, бұл NRF2 дамуын және протеазомдық деградацияны тудырады. Осы классикалық модуляциядан басқа, NRF2 сигналдық оқиғалармен терең реттеледі. Шынында да, әр түрлі киназалар NRF2-ді фосфорилаттайтын және реттейтін болып шықты. Мысалы, NRF2-ді митогенмен белсендірілген ақуыз киназаларымен (МАПК) фосфорлауға болады, дегенмен оның NRF2 белсенділігіне қосқан үлесі түсініксіз болып қалады [8], [9], [10], [11]. PKA киназа, сонымен қатар кейбір PKC изозимдері [12], CK2 [13] немесе Fyn [14] фосфорилат NRF2 оның тұрақтылығын өзгертеді. Біздің топтың бұған дейінгі жұмысы гликоген синтазы кинсе-3? (GSK-3?) NRF2-ді ядролық эксклюзия және протеазомдық деградация арқылы тежейді [15], [25], [26], [27], [28], [29], [30]. Молекулалық бөлшектер 4.1 бөлімінде талқыланады. Сонымен қатар, NRF2 басқа реттеу түрлеріне ұсынылады. Мысалы, CBP / p2 арқылы NRF300 ацетилдеуі оның белсенділігін арттырады [17], ал ол MiR153, miR27a, miR142-5p және miR144 [16] немесе NRF2 промоторы ішіндегі цитозин-гуанин (CG) аралдарының метилденуімен тежеледі. [18].
Протеиннің агрегаттарының NRF2 реттеу механизмдеріне әсері
Бұл бөлімде біз дұрыс емес ақуыздың жинақталуын NRF2 әсеріне қалай әсер етуі мүмкін екеніне баса назар аударамыз. Біріншіден, ақуызды жинақтау тотығу зақымымен тығыз байланысты болғанын ескеру керек. Шынында да, ақуыздың жинақталуы және агрегациясы митохондриядан және басқа көздерден (19) реактивті оттек түрінің (ROS) қалыпты өндірісін тудырады. Жоғарыда айтылғандай, ROS NRF1 шығарындыларының, тұрақтандырылуының және ядролық оқшаулаудың пайда болуына әкелетін KEAP2-ның редокс-сезімтал цистииндерін өзгертеді.
Протеинопатияларға қатысты NRF2-ге әсер етуі мүмкін реттелмеген сигналдық оқиғалардың мысалы GSK-3 гиперактивациясымен қамтамасыз етілген бе? б.з. GSK-3?, Сондай-ақ TAU киназа ретінде белгілі, осы микротүтікшемен байланысқан ақуыздың фосфорлануына қатысады, нәтижесінде оның агрегациясы, нейрофибриллярлы шатасулар түзілуі және аксональды тасымалдың тоқтауы ([20] -де қарастырылған). Екінші жағынан, GSK-3? жоғарыда айтылғандай NRF2 деңгейі мен белсенділігін күрт төмендетеді. Жалпы қабылданбағанымен, амилоид каскады улы А деп болжайды? олигомерлер GSK-3 ұлғайтады? TAU гиперфосфорлануымен және нейрон өлімімен бірге белсенділік [21], [22]. А-ны қалай түсіндіруге болатын әртүрлі модельдер бар? GSK3-ны қолдайды? белсенділік. Мысалы, А? инсулин рецепторымен байланысады және GSK-3-ті ұстап тұру үшін маңызды PI3K және AKT сигнализация жолдарын тежейді? оның Ser9 қалдықтарындағы N-терминалда фосфорлану арқылы инактивтелген [23]. Екінші жағынан, жасушадан тыс А? FNT рецепторларымен өзара әрекеттеседі, WNT сигнализациясын блоктайды [24] және қайтадан белсенді GSK-3 шығаруға әкеледі ?. Қысқаша айтқанда, А? жинақтау GSK-3 гиперактивациясының қалыптан тыс болуына алып келеді, осылайша NRF2 сәйкес реакциясын нашарлатады.
Келесі бөлімде талқыланғандай, қателенген ақуыздар NRF2 [31], [8], [9], [10], [11] -тарды реттейтін PERK және MAPK жүйелерін белсендіруге әкеледі. Сонымен қатар, CBP / p300 белсенділігімен дисчегулированная бірнеше протеинопатияларда [32] және ДНҚ метилирования жаһандық төмендеуі байқалды [33], сондықтан осы негізділігін анықтау үшін негіз береді NRF2 реттеу.
Біз және басқалар PD және AD науқастарында NRF2 протеинінің деңгейін жоғарылату және кейбір мақсаттары сияқты гемогенді оттегі, 1 (HMOX1), NADPH хинон оксидазы 1 (NQO1), p62 және т.б. иммуноблот және иммуногистохимия [34], [35], [36], [37], [38], [39] арқылы жасалады. Осы аурулардағы NRF2-нің реттелуі мидың гомеостатикалық құндылықтарды қалпына келтірудің сәтсіз әрекеті ретінде түсіндіріледі. Алайда, басқа зерттеу NRF2 негізінен AD гиппокампалы нейрондарының цитоплазмасында локализацияланғанын және мидағы [2] транскрипциясының NRF40 белсенділігін төмендететіндігін көрсетті. Бұл бақылаулардың дисбалансы нейродегенерацияның прогрессивті сатыларында NRF2-ды бақылайтын факторлардың өзгеруімен байланысты болуы мүмкін.
Протеостазға үш негізгі жүйе ықпал етеді: ақуыздың реакциясы (УПР), обицитин протеасомы жүйесі (UPS) және автофагия. Бұдан әрі NRF2-ты ақуыздың туындылы машиналары бар ақуыздық агрегаттармен белсендірілген авариялық сигналдарды байланыстыратын хаб ретінде қарастырамыз.
ЭР-де тотығатын протеиннің қатпарлануы бірқатар ерекше жолдармен қозғалады, олардың ішінде дисульфид доноры ретінде дисульфид-изомераза (PDI) және сульфгидрилоксидаза эндоплазмалық оксидоредуктин 1 (сүтқоректілерде ERO1? Және ERO1) бар. Қысқаша айтқанда, PDI өзінің цистеин аминқышқылдарының тотықсыздануы мен тотығуына байланысты ақуыздар ішіндегі цистеин қалдықтары арасындағы дисульфидтік байланыстардың түзілуін және бұзылуын катализдейді. ПДИ дисульфидті байланыстарды ПДИ-ге қайта енгізетін ERO1 үй шаруашылығының ферментінің әсерінен қайта өңделеді [41]. Молекулалық оттегі - бұл өндірілген дисульфидтің әр байланысы үшін стехиометриялық мөлшердегі сутегі асқын тотығын шығаратын ERO1 электронды терминал-акцепторы [42]. Пероксидазалар (PRX4) және глутатион пероксидазалары (GPX7 және GPX8) ER-да сутегі асқын тотығын төмендететін негізгі ферменттер болып табылады. Бұл оксидо-редуктивті жүйе дұрыс жұмыс істемегенде, қате ақуыздардың анормальды жинақталуы ER-да пайда болады және ER гомеостазын қалпына келтіру үшін цитоплазма мен ядроға белок реакциясы (UPR) деп аталатын сигналдар жиынтығы беріледі [43]. Эукариоттарда ER стрессін сезіну үшін мембранаға байланысты үш ақуыз анықталды: активтендіруші транскрипция факторы 6 (ATF6), ұйқы безі ER eIF2? киназа (PERK, сонымен қатар екі тізбекті РНҚ-активтендірілген ақуыз киназа тәрізді ER киназа) және инозитті қажет ететін киназа1 (IRE1). Әр сенсордың люминальды домені глюкозамен реттелетін ақуыз (GRP78 / BIP) деп аталатын 78 кДа шаперонмен байланысады. BIP қатпарланған ақуыздарды байланыстыру үшін ER стресс кезінде диссоциацияланады, бұл үш сенсордың активтенуіне әкеледі [44].
NRF2 және оның гомологтары NRF1, сонымен қатар антиоксиданттармен байланысты, UPR ядросына трансфекциясына қатысады. NRF1 жағдайда, бұл протеин ER мембранасында орналасқан және дегиклолиздену немесе бөлшектеу кезінде ядролық транслокацияға ұшырайды. Содан кейін UPR-ны қосу NRF1-ны өңдеуге және ядролық бөлімге алынған фрагменттің ядролық жинақталуына әкеледі. Алайда, NRF1 фрагментіндегі ARE бар гендерді транактивациялау мүмкіндігі әлі де талқылануда [45].
Glover-Cutter және әріптестері NRF2 ортопедиясын C. elegans, SKN-1, әртүрлі ER стресстерімен активтендіруді көрсетті. Артық SKN-1 өрнегі IRE1 немесе PERK құрт ортологтары [46] қоса алғанда, әртүрлі UPR медиаторларына тәуелді болды. ПЭК-жетіспейтін жасушаларда бұзылған ақуыз синтезі эндогендік пероксидтердің және кейінгі апоптоздардың жиналуына әкеледі [47]. PERK-дің осы пероксидтерден ER-ді қорғау үшін қолданатын эффекторы NRF2 болуы мүмкін, себебі PERX ферроматериалдары Ser2-де NRF40 фосфорланып, KEAP1 [31] арқылы оның бұзылуына жол бермейді. ASX1 индукциясы осы жолда IRE2 [1] TRAF48 арқылы жасалынған киназлы әрекеті арқылы рөл атқарады. NRF2-ті реттеудегі MAPK-дың рөлі әлі де қайшылықты болса да, жақында IRE1-TRAF2-ASK1-JNK жолының NRF2 [49] (1) функциясын белсендіруі мүмкін деп ұсынылған. Бір қызығы, C. elegans және адам жасушаларында жаңа деректер IRE1 киназының цистеиннің сульфенилдеуі оның белсендіру циклі кезінде IRE1-делдалдық UPR-ні тежейді және NRF38 арқылы басқарылатын p2 антиоксидантты жауапты бастайды. Деректер IRE1-дің ежелгі функциясы бар екенін көрсетеді, ол p38 және NRF2 [50] функциясын белсендіретін цитоплазмалық жөнелтуші болып табылады.
Сурет 1 UPR арқылы NRF2 реттеуі. Эндоплазмалық ретикулумның ішінде ашылған немесе қателенген ақуыздардың жиналуы ақуыздың (UPR) реакциясының бастамасы болуы мүмкін. Біріншіден, BIP шпарероны IRE1 ER және ERI сенсорларының ішінен шығарылған домендерден босатылған / қатпарланбаған ақуыздарды байланыстырады. Бұл олардың цитосолалық домендерінің димеризациясын және трансво-фосфорлануын қамтамасыз етеді. PERK активтендіруі NRF2 фосфорлануымен Ser40-те пайда болады, бұл ядроға NRF2 транслокациясына және мақсатты гендерді белсендіруге әкеледі. IRE1 активтендіруі TRAF2-ны жинауды, содан кейін ASK1 және JNK фосфорлануын және белсендіруді индуцирует. JNK фосфорилирлеу және NRF2 функциясын белсендірілгені туралы хабарланғандықтан, IRE1 белсендіру NRF2 белсенділігіне әкеледі деп ойлаған жөн.
ЖСЖ индукциясы бойынша көптеген зерттеулер ақуыз гликозилденуінің туникамицин ингибиторымен жүргізілді. NRF2 туникамициннің әсерінен пайда болған апоптотикалық жасушаның өлімінің алдын-алу үшін маңызды болып көрінеді [31] және оның осы жағдайда активтенуі KEAP1 аутофагиялық деградациясымен жүреді [51]. Тиісінше, NRF2 экспрессиясының shRNA арқылы тынышталуы? TC-6 жасушаларында, мирин инсулиномасы? -Желілік сызықта туникамицин туғызған цитотоксичность едәуір жоғарылап, про-апоптотикалық ER стресс маркерінің CHOP10 экспрессиясының жоғарылауына әкелді. Екінші жағынан, NRF2-ді 1,2-дитиол-3-тионмен (D3T) активациялау туникамициннің цитотоксикалығын төмендетіп, CHOP10 және PERK экспрессиясын әлсіретті [52]. Бір қызығы, туникамицинді жүйелік қолдануға ұсынылған иіс сезу нейрондары NOPF-ды CHOP, BIP, XBP2 сияқты басқа UPR мүшелерімен қатар арттырды [1]. Бұл нәтижелер in vivo зерттеулерге дейін кеңейтілді, өйткені егеуқұйрықтарға туникамициннің бүйірлік қарыншалық инфузиясы гиперпампуста PERK және NRF53 экспрессиясын тудырды, бұл когнитивтік жетіспеушіліктің жоғарылауымен, TAU фосфорлануының жоғарылауымен және A? 2 шөгінділерімен [42].
NRF2 ER физиологиясын жүргізу үшін негізгі гендерді жоғары деңгейде реттейді
Дисульфидті химияны ұстап тұру үшін ER люмені цитозолдан GSH мол қорын қажет етеді. NRF2 мидағы GSH метаболизмінің шешуші ферменттерін модуляциялайды, мысалы, цистин / глутамат тасымалы,? -Глутамат цистеин синтетаза (? -GS), глутамат-цистеин лигаза каталитикалық және модулятор суббірліктері (GCLC және GCLM), глутатион редуктазы (GR) глутатион пероксидаза (GPX) ([55] -де қарастырылған). NRF2-дің GSH-ті ұстап тұрудағы өзектілігі NRF2-дің фармакологиялық немесе генетикалық активтенуі GCLC / GCLM арқылы GSH синтезінің жоғарылауына әкеледі, ал NRF2-нокдаунмен осы ферменттердің экспрессиясын тежеп, зақымдалған заттардың жинақталуына әкеледі. UPR белсенділенуіне әкелетін ER ішіндегі белоктар [56].
C. elegans-де СКН-1 реттейтін UPR мақсатты гендерінің бірнеше компоненттері, соның ішінде Ire1, Xbp1 және Atf6. NRF2 сүтқоректілердегі бірнеше пероксидаза (PRX) және глутатион пероксидаза (GPX) гендерінің экспрессиясын жоғары реттейтініне қарамастан ([57] қарастырылған), тек GPX8 - KDEL іздеу сигналын сақтайтын, адал ER-локализацияланған ферменті [58]. GPX8 жоғалту UPR белсенділігін, ERO1? Алынған сутегі асқынының цитозолға ағып кетуін және жасушалардың өлуін тудырады. ERO1-ден алынған сутегі асқын тотығы? GPX8 және PRX4 келісілген әсерінен белсенділік ER-ден цитозолға дейін тарала алмайды [59]. Осыған байланысты, жабайы типтегі және NRF2-нөлдік тышқандар тінінен алынған РНҚ-ны қолдана отырып, антиоксидантты қорғаныс жолы-гендік экспрессиялық массивті талдау GPX8 экспрессиясының NRF2 болмаған кезде төмен реттелгендігін анықтады [60]. Осыған сәйкес, миелопролиферативті неоплазмалармен, полицитемиямен немесе миелофиброзбен, аурулармен ауыратын науқастардың сынамаларынан алынған транскриптомдық талдау, сонымен қатар тотығу стрессімен және төмен дәрежелі созылмалы қабынумен байланысты, бақылау субъектілерімен салыстырғанда NRF2 және GPX8 экспрессиясының төмен деңгейлерін көрсетеді [61]. Адамның миын қорғауға арнайы GPX8-ті қамтитын зерттеулер әлі жоқ, бірақ тышқандардағы транскриптоматикалық талдау паркинсондық токсин MPTP-ге жауап ретінде GPX8-нің компенсаторлық өсуін көрсетеді [62].
NRF2-ның нейродегенеративті аурулардағы UPR-дегі регуляциясына әсері
PDI ферменттерінің бұзылуы және UPR-ның созылмалы активтенуі кейіннен нейродегенерацияны бастамақ немесе жеделдетуі мүмкін. Аурулардан зардап шеккен нейрондар, нейродегенеративті аурулардың жануарлардың үлгілері, сондай-ақ адамнан кейінгі адамның ұлпалары бұл бұзылулардың көпшілігінде бірнеше UPR-маркерлерді жоғары деңгейде реттеуді дәлелдеді. PDR / UPR трассасының нейродежеративті аурулардағы өзгеруі [63] -те жақсы қарастырылды, бірақ мидан кейінгі үлгілердің келесі негізгі сәттері қарастырылуы керек. PDI деңгейлері науқас нейрондарда және AD және PD пациенттеріндегі Lewy органдарында [64], [65] -да жоғарылады. PDI және ERP57 ALS пациенттерінен CSF-де және CJD субъектілерінен [66], [67], [68] -тан алынған миында жаңартылған. AD, PD немесе ALS [1], [6], [69], [70] бар науқастардан BIP, PERK, IRE71 және ATF67 үлгілері жоғарылады. BIP, CHOP және XBP1 HD [72], [73] -тан кейінгі мидағы үлгілерде жоғарылайды. Сондай-ақ, CJD науқастарындағы [57] кортекс тіндерінде ERP94, GRP74 және BIP-нің реттеуі анықталды. Мұның барлығы мидағы паренхимадағы қателенген протеиндердің жинақталуы УДҚ-ның зиянды және созылмалы активациясына әкелетінін көрсетеді. Бір қызығы, жақында зерттеу басталғанға дейін NRF2 активациясын ПЕРК арқылы бастады. Зерттеу барысында авторлар NRF2 және UPR-дың ауытқушылықтарына байланысты тотығу стресстерінің өзгеруін талдады, бұл адамның перифериялық қан клеткаларын және әртүрлі ауру кезеңдерінде АЖ трансгенді тінтуір моделін қолдану арқылы АД патогенезінде ерте оқиғаларды құруы мүмкін. Күшті когнитивті бұзылулары бар адамдардан оқшауланған адамның перифериялық қандағы мононуклеарлы жасушаларында қышқылдықты жоғарылату және ұлғайту pSer40-NRF2 байқалды. Сонымен қатар, бұл клеткалардағы ER-кальций гомеостазының және ER-стресстің бұзылулары туралы жұмсақ когнитивті бұзылулары бар және жұмсақ AD [75] бар адамдардан хабарлады.
NRF2 және Ubiquitin Proteasome System (UPS) жүйесін өзара реттеу
UPS NRF2 ақуыз деңгейлерін өзгертеді
UPS зақымдалған немесе қателенген ақуыздардың тозуына қатысады және цитозольде және ядрода негізгі реттеуші молекулалардың деңгейлерін бақылайды. Бұл жүйенің орталық ядросы - 20S деп аталатын протеолитикалық белсенді комплексті бар көп молекулалық ферменттер. 20S ядросының протеасомы белоктарды нашарлатады, бірақ әр түрлі нормативтік белок комплекстеріне байланысу субстраттың ерекшелігі мен белсенділігін өзгертеді. Мысалы, 19S ядросына бір немесе екі 20S реттеуші бөлімшесі қосылып, 26S протеасомы болып табылады және оның [76], [77] протеиндеріне тәндігін өзгертеді. Протеасомалық деградацияда обицитиннің ковалентті байланыстары қажет. Убицитинді үш сатылы каскадының механизмі арқылы біріктіру. Алдымен, Ubququitin-белсендіретін E1 ферменті ATP-талап ететін реакцияда ubquitin белсендіреді. Содан кейін бір E2 ферменті (Убицитин-тасымалдаушы ақуыз немесе Убицитин-конъюганың ферменті) белсендірілген ubquitin E1-ден E3 деп аталатын Ubquitin-белоктар лигаз отбасының мүшесіне тікелей байланысты субстратқа тасымалдайды. Убицитинді-ақуыздың нақты тағдыры ubquitin тізбегінің табиғатына байланысты болса да, бұл процесс, әдетте, 26S протеасомы [78] арқылы ыдырауға әкеледі.
E3-лигоны KEAP1 - NRF2 ең жақсы белгілі ингибиторы. KEAP1 регламентінің механизмі NRF2 деңгейінің оксиданттың ауытқуларына қалай түзететінін түсіндіреді. Негізгі шарттарда жақында синтезделген NRF2 төмен (aspartate, lucine, glisin, DLG) және жоғары (glutamate, threonine, glisin, glutamate, ETGE) ұқсастығы бар екі аминқышқыл тізбегімен бір NRF1 молекуласын байланыстыратын KEAP2 гомодиаторы тәркіледі. KEAP1 құралдарымен өзара әрекеттесуі NRF2-ді CULLIN3 / RBX1 ақуыз кешеніне ұсынуға көмектеседі, нәтижесінде оның онтогенезі және кейінгі протеазомдық тозу. Дегенмен, KEAP1-дің redox-модификациясы CULLIN2 / RBX3 арқылы ұсынылған UPS-ге NRF1-ті ұсынуды кедергі келтіреді. Нәтижесінде жақында синтетизацияланған NRF2 KEAP1-тәуелді деградациядан құтылуға, ядрода жиналып, ARE құрамындағы гендерді [79], [80], [81], [82] гендерін белсендіреді.
E3-лигаза адаптері? -TrCP сонымен қатар GRK-2 әсерінен NRF3 фосфорлануымен байланысты сигналдық оқиғаларға қатысатын гомодимер болып табылады. Бұл киназа NRF2 (серпия, серин, глицин, изолейцин серині; DSGIS) белгілі бір сериндік қалдықтарын фосфорлайды, содан кейін? -TrCP арқылы танылатын және CULLIN1 / RBX1 комплексінің протеазомалық ыдырауы үшін белгіленетін деградациялық домен құру үшін. ГСК-3 фосфорланатын ерекше аминқышқылдарының идентификациясы? бұл дегронда Neh6 доменінің сайтқа бағытталған мутагенезі, 2D-гель электрофорезі [15], [26] және масс-спектроскопия [83] тіркесімі арқылы жүргізілді. Демек, ГСК-3 тежелуі? GSK-3 изоформаларына қарсы жоғары селективті дәрілермен немесе сиРНК-мен NRF2 ақуыз деңгейінің жоғарылауына әкелді. Ұқсас нәтижелер SiRNA-мен? -TrCP изоформаларының 1 және 2-ге қатысты болды, GSK-2-тен кейін NRF3 тұрақтануы? тежеу KEAP1 жетіспейтін тышқан эмбрионының фибробласттарында және KEAP2-ге тәуелділігі жоғары байланыстыру үшін ETGE сыни қалдықтары жоқ эктопиялық экспрессияланған NRF1 жою мутантында орын алды, бұл әрі қарай KEAP1 тәуелсіз реттелуін көрсетеді.
Нейродегенеративті аурулар аясында біз UPS арқылы NRF2 модуляциясын екі түрлі жолмен елестете аламыз. Бір жағынан, KEAP1 жүйесі ақуыздың дұрыс жиналмауынан пайда болған тотығу-тотықсыздану тепе-теңдігін сезінеді, ал GSK-3 /? - TrCP осі протеостаздың жоғалуымен өзгертілген сигналдық трансдукцияның белсенді қатысушысы ретінде әрекет етеді (2-сурет).
Сурет 2 UPS NRF2 деңгейлерін тығыз басқарады. Гомеостатикалық жағдайда NRF2 деңгейінің төмендігі KEAP3 және? -TrCP E1 лигаза адаптерлерінің әсерінен сақталады. Сол жақта NRF2 төмен (DLG) және жоғары (ETGE) жақындық мотивтері арқылы KEAP1 гомодимерінің Kelch домендерімен байланысады. BTE домені арқылы KEAP1 бір мезгілде CULLIN3 / RBX1 кешенімен байланысады, бұл NRF2-дің барлық жерде орналасуына және 26 S протеазомасының ыдырауына мүмкіндік береді. Сонымен қатар, GSK-3? NRF335-нің Ser338 және Ser2 қалдықтарын фосфорлайды, содан кейін ubiquitin ligase адаптері? -TrCP арқылы танылатын және CULLIN3 / RBX1 комплексімен протеазоманың деградациясы үшін белгіленген деградациялық доменді (DpSGIpSL) құру үшін. Оң жақта, реактивті оттегі түрлеріне немесе электрофилдерге әсер еткенде KEAP1-дегі маңызды Cys қалдықтары өзгертіліп, KEAP1 NRF2 немесе CULLIN3 / RBX1-мен тиімді әрекеттесе алмайды, содан кейін бұл транскрипция коэффициенті ARE-гендерге қатысты жартылай шығарылу кезеңін және транскрипциялық белсенділігін арттырады. G3-тің тежелуіне әкелетін сигналдық жолдар, Ser9-дағы осындай АКТ фосфорлануы NRF2-нің протеазоманың, мақсатты гендердің жиналуы мен индукциясының нашарлауына әкеледі.
NRF2 протеасомды қосалқы элементтерді транскрипциялық бақылау арқылы UPS қызметін көбейтеді
NRF2 бірнеше протеазомалық суббірліктердің экспрессиясын реттейді, осылайша жасушаны улы протеиндердің жиналуынан қорғайды. NRF2 индукторы D2T көмегімен орнатылған бауыр РНҚ-сынан алынған кең микроарриздік анализге сәйкес, протеазомамен және барлығымен байланысты жиырма ген NRF3 арқылы реттелетін көрінеді. Артқы зерттеу кезінде дәл сол авторлар 84S протеазомасының көптеген суббірліктерінің экспрессиясы D26T-мен өңделген тышқандардан бауырда үш есеге дейін жоғарылағанын дәлелдеді. Суббірлік ақуыз деңгейі және протеазоманың белсенділігі үйлесімді түрде жоғарылаған. Алайда NRF3 транскрипциясы факторы бұзылған тышқандарда индукция байқалмады. PSMB2 (5S) протеазома суббірлігінің промотор белсенділігі NRF20 шамадан тыс экспрессиясымен немесе тінтуір эмбриональды фибробласттарындағы активаторлармен емдеумен жоғарылаған және PSMB2 проксимальды промоторында AREs анықталған [5]. NRF85-ді фармакологиялық активтендіру тек функционалды NRF2 бар адамның кәрілік емес фибробласттарындағы протеазомалық суб-бірліктердің (PSMA3, PSMA6, PSMB1 және PSMB5) экспрессия деңгейінің жоғарылауына алып келді [2]. Тотығу стрессіне бейімделу кезінде NRF86 активациясы PSMB2 (1S) және PA20 жоғары экспрессиясына әкеледі? суббірліктер (немесе S28, протеазома реттегіші) [11]. Сонымен қатар, адамның эмбриондық бағаналы жасушаларының нәтижелері бойынша NRF87 протеазомалық жетілу протеинінің (POMP) экспрессиясын басқаратындығы анықталды, бұл өз кезегінде адамның өзін-өзі жаңартатын эмбриондық бағаналы жасушалардың көбеюін, үш ұрық қабатын дифференциациялауды және жасушалық қайта бағдарламалауды модуляциялайды [ 2]. Бұл зерттеулер NRF88 UPS-тің негізгі компоненттерінің экспрессиясын реттейтіндігін көрсетеді, сондықтан басқа жағдайда улы болатын белоктардың тазартылуына белсенді ықпал етеді.
Нейродежеративті аурулардағы NRF2-UPS осі
ИБП-ның нейродегенеративті аурулардағы рөлі қарқынды пікірталас алаңы болып табылады. Алғашқы зерттеулерде бірнеше нейродегенеративті аурулардан зардап шеккен науқастарда адамның асқазанында протеазомдық белсенділіктің төмендегені туралы хабарлады. Алайда, in vitro және in vivo тәсілдерде қолданылатын басқа зерттеулерде протеасомдық белсенділіктің өзгермегендігі немесе тіпті жоғарылауы анықталды (89-де қаралды). Бұл сәйкессіздіктің мүмкін болатын түсіндірмесі - бұл NRF2-мақсаттары үшін ұсынылғандай, UPS компоненттерінің деңгейлері аурудың дамуын және әр түрлі ми аймақтарында өзгеруі мүмкін.
Осы қарама-қайшылыққа қарамастан, ARE құрамындағы протеасомдық гендердің жоғары реттелуі мидағы уытты протеиндердің тазалығын арттыру арқылы ИБП-ны күшейтеді. Шынында да, NRF1 абляциясы, сондай-ақ антиоксиданттың реакциясының модуляторы нейрондық жасушаларда протеасомдық белсенділікке және нейродегенерацияға әкеледі. Chromatin иммунопреципитация эксперименттері және транскрипциялық талдау PSMB6 NRF1 арқылы реттелетінін көрсетті. Сонымен қатар, гендік өрнекті профильдеу NRF1-тың нейрондардағы протеасомдық гендердің негізгі транскрипциялық реттеушісі ретінде анықтауға әкелді, бұл NRF1-те наразылық нейродегенеративті аурулардың [90] патогенезіне ықпал етуі мүмкін екенін көрсетеді. Бір қызығы, NRF1 және TCF11 деп аталатын оның ұзақ изооформасы ARE құрамында протеазомдық гендерді кері байланыс циклындағы протеазомдық ингибицияны азайту үшін көрсетілген [91], [92].
NRF2 болсақ, NRF2, RPT6 (19 S) және PSMB5 (20 S) деңгейлерінің арасында нейротоксин паракуатымен [1] емделген DJ-93-жетпейтін тышқандардың ортасында корреляция бар. Сонымен қатар, табиғи сульфарфан (SFN) құрамында NRF2 неғұрлым берік бейнесі ИБП-ның аса маңызды модуляторы болып табылады. Нейро-ХНУМХА жасушаларының миокард нейробластомасы бар In vitro эксперименттері протеасомның каталитикалық суб-бөліктерінің күшейтілген көрінісін және SFN-ке жауап ретінде оның пептидаздық әрекеттерін дәлелдеді. Бұл препарат протеинді функцияға байланысты [2] сутектік пероксидтен цитотоксичность және ақуызды тотығудан жасушаларды қорғайды. Бұдан басқа, Лю және әріптестер Миға SFN-ке жауап ретінде ИБП-ның қызметін бақылау үшін журналистердің тышқанымен жұмыс істеді. Бұл тышқандар UPS (GFPu) арқылы тез бұзылуына ықпал ететін конструктивті деградация сигналына ұшыраған жасыл флуоресцентті ақуызды (GFP) білдіреді. Церебральды кортексте SFN GPPu деңгейін химотрипсин тәрізді (PSMB94), caspase-like (PSMB5), және трипсин тәрізді (PSMB2) 1 S протеазомының параллель жоғарылауымен төмендетті. Сонымен қатар, SFN бар Хантингтоннан алынған жасушаларды емдеу NRF20 активтендіруі мГт деградациясының және мутекті цитотоксичность [2] деңгейін азайтты. SFN әрекетінің негізгі механизмі NRF95 [2] индукция арқылы жүзеге асырылады. NRF96-тің ерекше үлесі NRF2-нөл жүйелерін әрі қарай зерттеуде қолдануға арналған.
NRF2 және Macroautophagy арасындағы функционалдық қосылу
NRF2 ақуыз деңгейлері адаптердің P62 протеині арқылы өзгертіледі
Автофазия лизосомалардың ішінде цитозолиттік компоненттердің тозуына қатысты. Бұл процесс ұзақ уақыт өмір сүретін және қателенген ақуыздардың, сондай-ақ зақымдалған органеллдердің тазалануы үшін қолданылады. NRF2 мен autophagy арасындағы тікелей байланыс алғаш рет SQSTM62 [1], [97], [98], [99], [100] деп аталатын адаптер ақы p101-мен байланысты. Бұл ақуыз протеазомдық және лизосомалық деградациялау машиналарына белдеулерді ұйытып, белоктарды агрегацияға дейін бұзады. P62 - Ubquitine-associated (UBA) доменді, олимпиадалық рецепторлар LC3 арқылы автофагосомалық мембранамен интеграциялану үшін, Ubiquitinated протеиндерге және LC3-өзара әрекеттесетін аймаққа (LIR) байланысады.
NRF62 және оның мақсатты гендерінің p2-индукциясы туралы алғаш рет 2007 жылы хабарланғанымен [102], молекулалық механизм оның KEAP1 [103], [98], [99], [100] өзара әрекеттесуі ашылғанға дейін толық түсінілмеген. ], [101]. Komatsu және оның әріптестері p1 мен KRR62 сияқты негізгі беткі қалтада KEAP1 байланыстыратын және NRF2 ішіндегі ETGE мотивіне ұқсас байланыстырушы қабықпен p2 және NRF62 арасындағы бәсекелестікті болжайтын KEAP2 өзара әрекеттесетін аймақты (KIR) анықтады. P351 (62-DPSTGE-349) ішіндегі KIR мотивіндегі Ser354 фосфорлануы оның NEF1 байланыстыруымен бәсекелесетін және оның мақсатты гендерінің жинақталуына және транскрипциялық активтенуіне мүмкіндік беретін KEAP2-ге жақындығын арттырды [98], [99]. Шындығында, p62 шамадан тыс экспрессия NRF2 барлық жерде азайып, нәтижесінде оның тұрақтандырылуына, сондай-ақ оның мақсатты гендерінің индукциясына әкелді [104]. Кейбір киназаларға p62 фосфорлануға қатысуға ұсыныс жасалды. Рапамицин кешенінің 1 (mTORC1) сүтқоректілерінің мақсаты болуы мүмкін, өйткені mTOR ингибиторы рапамицинмен емдеу p62 фосфорлануын және арсенитпен емдеу кезінде KEAP1-нің төмен реттелуін басады. Жақында TGF -? - активтендірілген киназа 1 (TAK1) KEAP62 деградациясын және NRF1-реттелуін күшейтетін p2 фосфорилденуі мүмкін екендігі дәлелденді. Зерттеудің авторлары бұл тұрақты күйдегі жасушалық редокстазды реттеу әдісін ұсынады, өйткені TAK1 жетіспеушілігі әр түрлі тышқан тіндерінде экзогендік тотықтырғыш болмаған кезде ROS-ны реттейді, NRF2 ақуыз деңгейінің төмендеуімен қатар ].
UBA домені болмаған p62 құрамы әлі де KEAP1-ты байланыстыруға қабілетті, бұл өзара әрекеттесу KHAP1 [101] -қа тәуелді емес дегенді білдіреді. Дрозофила меланогастеріндегі Ref (62) деп аталған p2 гомологы KIR мотивтерін қамтымайды және DmKEAP1-мен тікелей өзара әрекеттеспейді, бірақ ол UBA доменінің көмегімен Ubququitinated DmKEAP1-ке қосыла алады. Сонымен қатар, DmKEAP1 тікелей Atg8 (сүтқоректілер LC3 гомологтарымен) өзара әрекеттесе алады. KEAP1 тапшылығы Atg8 және autophagy индукциясына NRF2 ортолог CncC тәуелді және TFEB / MITF [106] тәуелсіз нәтиже береді. NRF2 мен автофаги арасындағы байланыс өзінің функционалдық маңыздылығымен ерекшеленетін болса да сақталады.
P2 арқылы NRF62 индукциясы - KEAP1 мен KEAP1-ты лизосомада байланыстыратын бәсекелестіктің нәтижесі. SiRNA бар p62 дыбысын өшіру NRF1 және оның мақсатты гендерінің [2] азаюымен параллель KEAP101 жартысын қосады. Келісімде p62 өрнегінің абляциясы жабайы түрдегі тышқанмен салыстырғанда KEAP1 деңгейін жоғарылатты. Өте маңызды, KEAP1 деңгейлеріндегі протеазомдық ингибиторлар әсерін тигізбеді, бірақ аштықтан кейінгі ауофагия кезінде [107] азайды. Шын мәнінде, KEAP1 p62 және LC3 [99], [100], [103] көмегімен безендірілген аутоафагиялық весикулалардағы сүтқоректілердің жасушаларында болады. Осы деректердің барлығы KEAP1 - макрофотозды техниканың субстраты екендігін көрсетеді, бірақ бұл мәселені кейбір даулы нәтижелердің болуына байланысты егжей-тегжейлі талдау қажет. KEAP1 белок деңгейлері Atg7-null тышқандарында ұлғайтылды, макроэтофаги [107] негізгі эффекторы, бірақ макроэтоофагияның torin1, E64 / pepstatin немесе bafilomycin фармакологиялық ингибирлеуі KEAP1 [107], [100] жинақталмады. Тұтастай алғанда, бұл нәтижелер P62 деңгейін KEAP1 секциясының автофагиялық вакуолдарға көбейтіп, NRF1 активтендіруіне мүмкіндік беретін аутофагиялық ыдыраудың KEAP2 нәтижелеріне әкелуі мүмкін (3 сур.). Екі түрлі зерттеудің деректері бойынша, SENTINS сульфинді қышқылын қалпына келтіреді, осы тұрғыда маңызды рөл атқарады. SESTRIN 2 p62, KEAP1 және RBX1-пен өзара әрекеттеседі және мақсатты гендердің KEAP62 және NRF1 активтілуінің p2-тәуелді деградациясына көмектеседі [108]. Басқа зерттеу SESTRIN 2 UX1 және p62-пен өзара әрекеттесетіндігін көрсетті, ол Ser62-дағы p403 фосфорлануын арттырады, соның арқасында KEAP1 [109] жүктің белоктарының деградациясы жеңілдейді.
Сурет 3 NRF2 деңгейлері адаптер ақуызы p62 арқылы реттеледі. Ser351-дің p62 (349-DPSTGE-354) KIR мотивтерінде mTORC1, TAK1 немесе басқа киназдардағы ферменттелуі FNF1-те ETGE мотивіне ұқсас болуына байланысты KEAP2-ге байланыстыру үшін жоғары жақындығымен байланысты. Соның салдарынан, фосфорилирленген p62 NRF2-ты ауыстырады және KEAP1-ді байланыстырады. P62 ішіндегі LIR мотиві autofagosomal мембранадағы LC3-пен өзара әрекеттесуге мүмкіндік береді, сондықтан p62-KEAP1 кешені лизосомада бұзылады. NRF2 нәтижесінде NRF62 жинақталып, ядроға ауысып, ARE құрамындағы гендерді, соның ішінде p2 транскрипциясын көбейте алады. Бұл реттеуші механизм NRF1 әсерін қамтамасыз етеді, себебі NRF2 белсенділігін ингибирлеу үшін KEAPXNUMX жаңадан синтезделуі керек.
NRF2 арқылы макрофотозды гендердің модуляциясы
NRF2 макроэтоофагия үшін тиісті гендердің өрнегін реттейді, сондай-ақ UPR және ИБП үшін қолданылады. Алғашқы дәлелдер p62 экспрессиясының электрофильдер, ROS және азот оксидтері [110], [111], [112] әсерінен туындаған зерттеулерден алынған. Индукция механизмі бірнеше жыл өткеннен кейін p62 өзінің гендік промоутерінде [99] функционалды ARE бар екендігі туралы анықталған. Жақында өткізілген зерттеуде биоинформатикалық талдау мен ChIP талдауынан кейін бірнеше басқа функционалды АЖ-ні анықтады және тексерді. Сонымен қатар, Nrf2-нокаут тышқандарынан тышқанның эмбрионды фибробласттары мен кортикальды нейроны NRF62 экспрессивті лентивируспен құтқарылуы мүмкін p2 сөйлемшесін азайтты. Сол сияқты, NRF2 жетіспеушілігі зақымдалған нейрондарда тышқанның гиппокампусынан [62] алынған p36 деңгейлерін төмендетті. Сондықтан, NRF2 активтендіруі p62 деңгейін жоғарылатады, бұл KEAP1 деградациясына әкеледі және оң нәтиже беру циклі бойынша NRF2 тұрақтылығын қолдайды. NRF2 индукциясының бұл канондық емес механизмі геннің өрнегіндегі өзгерістерді талап етеді және созылмалы жасушалық кернеуге жауапты болуы мүмкін.
NDP52 жүкті тану белок NRF2 арқылы транскрипциялық түрде реттеледі. NDP52, p62-ге ұқсас жолмен жұмыс істейді, ол LQ3-мен LYL доменімен өзара әрекеттеседі, ол либосозомдарда бұзылады. Ndp52 промокаторының ДНҚ тізбегіндегі бес болжамды ARE табылды. Олардың үшеуі NRF2-делдалдық Ndp52 транскрипциясындағы [113] транскрипциясы үшін әртүрлі мутант конструкциялары мен ChIP талдауларымен анықталды. Оның үстіне, Nrp52 мРНҚ деңгейлері Nrf2-нокаутқа арналған тышқандардағы гиппокампада азайған. Осы тізбектердің бірі NRF2-реттелген ARE [36] ретінде тәуелсіз зерттеуде де жарамды болды.
Дегенмен, автофагияның модуляциясындағы NRF2 рөлі осы екі жүк тану белоктарының индукциясымен шектелмейді. NRF2-тың қосымша автофагиямен байланысты гендерді модуляциясында тереңірек түсіну үшін біздің топ NRF1-реттелетін ARE-лерді байланыстыратын екі протеин, MAFK және BACH2 үшін хроматиннің иммунопреципидатының ENCODE дерекқорын көрсетті. JASPAR-ның консенсусы ARE жүйесінен алынған сценарийді пайдалану арқылы көптеген автофазалық гендерде бірнеше болжамды АЖ-н анықтадық. Осы тізбектердің он екісі NRF2 регламенттелген AREs ретінде Nrf2-нокаутқа тышқандарындағы фибробласттардың тінтуір эмбрионында азаяды, бірақ NRF2-экспонатталған лентивирус арқылы қалпына келтірілген тоғыз ауто-фазалық гендер ретінде бекітілді. Біздің зерттеу NRF2 автофагиялық процестің әртүрлі кезеңдеріне қатысатын кейбір гендердің экспрессиясын белсендірді, соның ішінде автофагияның бастамасы (ULK1), жүкті тану (p62 және NDP52), автофагосома қалыптастыру (ATG4D, ATG7 және GABARAPL1), ұзарту (ATG2B және ATG5) ), сондай-ақ автолисосомалық тазарту (ATG4D). Демек, NRF2 жоқ болған кезде сутегі асқын тотығына жауап ретінде автофагия ағыны бұзылған [36].
Ақаулы автофагия бірнеше нейродегенеративті ауруларда маңызды рөл атқарады [114] және ауофазияның абляциясы тышқандардағы [115], [116] нейродегенерациясына әкеледі. Atg7-нокаут тышқандарында autophagy жетіспеушілігі ubquitin-оң инклюзивті органдарда p62 жинақталуының нәтижесін көрсетті. KEAP1 осы инклюзивті органдарда секвенирленген, NRF2 тұрақтандыру және мақсатты гендердің индукциясы [103]. Маңыздысы бойынша, P62-нің белокты жинақталған ақуыздармен бірге, AD, PD және ALS [117] қоса алғанда нейродегенеративті ауруларда шамадан тыс жинақталуы анықталды. Іс жүзінде жоғары деңгейдегі APP немесе TAU пациенттерін білдіретін нейрондар p62 және ядролық NRF2-ты білдірді, бұл олардың аутогенді агрегаттарды автофагия арқылы [36] бұзу әрекетін болжайды.
NRF2 тапшылығы АД аясында ақуыздың агрегациясын күшейтеді. Іс жүзінде фосфорланған және саркозилде ерімейтін ТАУ деңгейінің жоғарылауы Nrf2-нокаут тышқандарында кездеседі, дегенмен жабайы типтегі фонмен салыстырғанда киназа мен фосфатаза белсенділігінің айырмашылығы анықталмады [113]. Маңыздысы, NDP52 мириндік нейрондарда ТАУ-мен бірге локализацияланғаны және фосфо-ТАУ мен NDP52 арасындағы тікелей өзара әрекеттесуі тышқандарда да, AD үлгілерінде де иммунопреципитация эксперименттерімен көрсетілген, бұл оның ТАУ деградациясындағы рөлін көрсетті. Бір қызығы, NDP52, p62 немесе NRF2 нейрондарда тынышталуы фосфо-TAU деңгейінің жоғарылауына әкелді [113], [118]. Сонымен қатар, NRF1 болмаған кезде APP / PS9? E2 тышқандарының гиппокампасында интранеуральды APP агрегаттары табылды. Бұл өзгертілген аутофагия маркерлерімен, соның ішінде фосфо-mTOR / mTOR және фосфо-p70S6k / p70S6k коэффициенттерімен (аутофагияның тежелуін көрсететін), D-катепсинге дейінгі деңгейлердің жоғарылауымен және мультивискулярлы денелердің үлкен санымен байланысты болды [119]. Адамның APP (V717I) және TAU (P301L) экспрессивті тышқандарында NRF2 жетіспеушілігі ерімейтін фракциядағы жалпы және фосфо-ТАУ деңгейінің жоғарылауына және p62, NDP52, ULK1 нейрондық деңгейлерінің төмендеуімен интранейрональды APP агрегаттарының жоғарылауына әкелді, ATG5 және GABARAPL1. P62 адаптері мен APP немесе TAU адаптері арасындағы бірлескен оқшаулау NRF2 болмаған кезде азайды [36]. Жалпы алғанда, бұл нәтижелер NRF2-нің нейрондық аутофагиядағы маңыздылығын көрсетеді.
Протеостазды модуляциялау үшін үйлесімді түрде түрлі транскрипция факторлары
Тұрақты күй жағдайында протеостаз протеин-ақуызмен өзара әрекеттесу арқылы және жылдам жауап алудан кейінгі трансволюциялық өзгерістермен бақыланады. Дегенмен, ұялы бейімдеу UPR, UPS және autophagy гендерінің транскрипциялық реттелуін талап етеді. Нерв клеткаларының төменгі дәрежелі уытты қорлауға, соның ішінде тотығу және протеотоксикалық стресске ұшырататынын ескере отырып, транскрипциональды модуляция арқылы туындаған протеостаздың күшейтуі мидың деградациясының алдын алуға көмектесуі мүмкін.
UPR жағдайда, үш қолдың әрқайсысының белсенуі белгілі бір гендердің транскрипционды индукциясына әкеледі (43-де қарастырылған). Мысалы, ATF6 фрагменті (ATF6f) ER-стресс жауап элементтеріне (ERSE) байланыстырады және XBPI, BIP және CHOP қоса бірнеше геннің өрнегін индуцирует. Сонымен қатар, PERK сигналы HF4 транскрипция коэффициентінің белсендірілуіне әкеледі, ол бірнеше UPR-байланысты гендердің және басқалардың NRF2 мақсатты генін Hmox1 және p62 гендеріне жатқызады. Соңында, IRE1 белсендіру белсенді транскрипция коэффициентін қалыптастырады, XBP1 (XBP1s), протеинді жинауға қатысатын ақуыздарды кодтайтын гендердің транскрипциясын басқарады.
Екінші жағынан, NRF1 мидағы протеазомальды геннің өрнегі үшін қажет болғандықтан, Nrf1-нокаутқа тышқандар 20S ядросының әртүрлі бөлімдерін кодтайтын гендердің экспрессиясын азайтты, сондай-ақ, 19S реттеуші комплексі бұзылған протеазомдық функциямен бірге [90 ]. NRF1 және NRF2 екеуі де мақсатты гендердің промоутерлік аймақтарындағы ARE жүйесімен байланысады, бұл олардың реттелетін тетіктерінде және ұялы оқшаулауында [120] айырмашылығы бар болса да, олардың транскрипциялық әрекеттерге сәйкес келетіндігін көрсетеді.
Forkhead қорғаныс қорабының транзитивтік факторлары O (FOXO) отбасылық бақылауды бірнеше аутоафагиямен байланысты гендердің экспрессиясын бақылайды. NRF2-пен болатын жағдайға ұқсас, FOXO мүшелерінің белсенділігін реттеудің бірнеше топтары бар, олар қоректік немесе тотығу стресс кезінде пайда болуы мүмкін [121]. Ақырында, лизосомалық биогенездің негізгі реттеушісі деп саналатын TFEB транскрипция коэффициенті қоректік кернеулер жағдайында автофагияны реттеуде шешуші рөл атқарады. Осылайша, mTORC1 ингибировкасы TFEB ядролық транслокациясына және автофагия гендерінің экспрессиясын [122] әкеледі.
Тұтастай алғанда, осы машиналардың түрлі транскрипциялық реттегіштерінің болуы, сонымен қатар, әртүрлі жағдайда протеостазды қамтамасыз етуі мүмкін кросстелк пен ішінара артық тетіктерді ұсынады. Тиісінше, NRF2 тотығу стрессінің жоғары деңгейін қолдайтын маталардағы маңызды рөл болуы мүмкін. Мысалы, тотығу стресс-индуцирленген NRF2 аштық жағдайында TFEB үшін табылған нәрсеге ұқсас, автофагияны транскриптті түрде реттеуге арналған қоректік заттарға бай жағдайлар кезінде жұмыс істей алады. Оның үстіне, ми жұмыс істейді, негізінен қоректік заттарға бай жағдайларда, NRF2-ды нейрондарда автофазияны белсендірудің тиісті механизмі ретінде көрсетеді.
Протеинопатиялардағы NRF2 үшін перспективті терапиялық потенциал
Соңғы бірнеше жылда NRF2 белсенділігінде UPR, UPS және автофагияның реттелетін рөлдерін, сондай-ақ осы үш жүйенің құрамдас бөліктерінің өзара NRF2 арқылы транскрипциясын білу үлкен жетістікке қол жеткізді. Сондықтан NRF2-ды нейродегенеративті аурулардағы протеинді тазартудың маңызды реттегіші ретінде пайдаланудың жаңа терапиялық мүмкіндіктері пайда болуы мүмкін.
Дегенмен, қалған негізгі сұрақ - мидағы NRF2 деңгейін жоғарылату пайдалы немесе зиянды ма. Эпидемиологиялық деректерді талдау ішінара жауап бере алады, өйткені ол NFE2L2 генінің жоғары полиморфты екенін көрсетеді және оның промоторының реттеуші аймағында табылған кейбір жалғыз нуклеотидтік полиморфизмдер популяция деңгейінде және кейбір гаплотиптерде ген экспрессиясының «физиологиялық» өзгермелілігін қамтамасыз ете алады. AD, PD немесе ALS қаупінің төмендеуімен және/немесе кешігуімен байланысты болды [123]. Сонымен қатар, Хейс және әріптестері [124] талқылағандай, NRF2 әсері U-тәрізді жауапқа ие болуы мүмкін, яғни тым төмен NRF2 деңгейлері цитопротекцияның жоғалуына және стресс факторларына сезімталдықтың жоғарылауына әкелуі мүмкін, ал NRF2 тым көп болуы гомеостаздық тепе-теңдікті бұзуы мүмкін. ақуыздың қате қатпарлануын және агрегациясын қамтамасыз ететін редукциялық сценарий (редукциялық стресс). Мидағы төмен NRF2 деңгейлері патологиялық жағдайларда пайдаға жету үшін шамалы жоғарылату жеткілікті болуы мүмкін деген идеяны қолдайды. Шын мәнінде, ақуыз клиренсінің фармакологиялық NRF2-делдалдық белсендіруінің қорғаныш рөлі әртүрлі нейродегенерация жасушаларының мәдениетінде және in vivo үлгілерінде көрсетілген.
SFN - протеазомдық және аутофагиялық гендердің экспрессиясын тудыратын NRF2 фармакологиялық активаторы [95], [36]. Бір қызығы, Джо және оның әріптестері SFN фосфорланған ТАУ деңгейін төмендетіп, Беклин-1 және LC3-II жоғарылағанын көрсетті, демек, NRF2 активациясы бұл улы протеиннің аутофагия арқылы ыдырауын жеңілдетуі мүмкін [113]. Сонымен қатар, mHtt деградациясы SFN көмегімен күшейтілді және бұл MG132 қолдану арқылы қалпына келтірілді, бұл осы улы протеиннің протеазомальды деградациясын көрсетеді [95]. Органикалық флавоноидты фисетинмен бірге фосфордың және ерімейтін-ТАУ-дың аутофагиямен ыдырауы туралы хабарланды. Бұл қосылыс TFEB-нің де, NRF2-нің де, оның мақсатты гендерімен бірге активациясын және ядролық транслокациясын бір мезгілде алға жылжыту арқылы аутофагия тудырды. Бұл жауап TFEB немесе NRF2 тынышталуының алдын алды [125]. Ботт және оның әріптестері бір мезгілде NRF2, NRF1 және HSF1 активаторының белоктар мен омыртқаның бұлшықет атрофиясындағы белоктың уыттылығына, полиглутаминді кодтайтын CAG қайталануының кеңеюінен туындаған нейродегенеративті бұзылысқа пайдалы әсерлері туралы хабарлады [126]. Нейродегенеративті бұзылыстарды емдеуге арналған NRF2 активациясының мүмкіндігі көптеген склерозды емдеу үшін NRF12 индукторының диметил фумаратының (DMF) ауызша формуласы болып табылатын BG-2 мақұлдауымен дәлелденді [127], [128]. Қатты қабыну компоненті бар аутоиммунды аурулармен DMF-дің жетістігі нейродегенеративті аурулар осы препаратты қайта орналастырудан пайда табуы мүмкін. Жақында ПД-синуклеинопатия моделін клиникаға дейінгі зерттеу барысында DMF ішінара аутофагия индукциясына байланысты нейропротекторлық болып шықты [129]. NRF2-нің нейродегенерацияға пайдалы әсерлері туралы есептер, бірақ оның ақуыз клиренсіне әсеріне назар аудармайтын зерттеулер одан да көп (толық шолу үшін [7] қараңыз). Бұл өте өзекті, өйткені ол NRF2-де бір рет соққыға бағытталуы мүмкін көптеген зақымдайтын процестерді, сонымен қатар тотығу стрессін, нейроинфламмацияны немесе митохондриялық дисфункцияны қамтиды. Алайда, NRF2-дің фармакологиялық активтендірілуі мидағы улы белоктардың деградациясын жеңілдететін жарамды стратегия бола ма, жоқ па, соны анықтау үшін болашақ жұмыс қажет болады.
Бұрын түсіндіргендей, ГСК-3 күшейе түсті ме? нейродегенеративті ауруларда белсенділік туралы хабарланды және NRF2 төмендеуі зиянды нәтижеге ішінара жауапты болуы мүмкін деген болжам жасалды. Осы патологиялық жағдайларда GSK-3 ингибиторлары NRF2 деңгейін және протеостазды арттыру үшін де ынтымақтаса алады. GSK-3 ингибиторларының пайдалы әсерлері нейродегенерацияның әртүрлі модельдерінде хабарланған және қызықтыратыны, GSK-3 репрессиясы улы белоктардың деңгейін төмендететіні көрсетілген [130], [131], [132], [133]. Протеостазды дамытатын гендердің GSK-3 тежелуі мен NRF2-транскрипциялық реттелуі арасындағы тікелей байланыстар әлі байқалмағанымен, GSK-3 белсенділігінің төмен реттелуі NRF2 деңгейінің жоғарылауына алып келеді, бұл ақырында күшейтіледі деп болжау орынды. протеостаз.
NRF2 транскрипциялық белсенділігі, сондай-ақ протеостазды ұстап тұру үшін жасушалық қабілет жасына қарай төмендейді, бұл нейродегенеративті аурулардың дамуының негізгі қауіп факторы. NRF2-ны күшейту және, демек, протеостаз, кем дегенде, ақуыз агрегаттарының жиналуын және нейродегенерацияны кешіктіреді деп ойлау орынды. Шынында да, адамның қартайған фибробласттарын 18? -Глициретин қышқылымен (18? -GA) тритерпеноидпен емдеу NRF2 белсендірілуіне ықпал етіп, протеазомалық индукцияға және өмірдің ұзаруына әкелді. Бұл зерттеу NRF2-ді фармакологиялық активтендіру өмірдің соңғы кезеңінде де мүмкін болатындығын көрсетеді [86]. Сонымен қатар, кейінгі зерттеу көрсеткендей, бұл қосылыс SKN-1 мен C. elegans протеазомаларын белсендіруге сәйкес нематод модельдерінде AD прогрессиясына тиімді әсер етеді [134].
Барлық қарастырылған, NRF2 арқылы протеостазбен байланысты гендерді индукциялау әртүрлі белоктардың патологиясында пайдалы болып көрінеді.
Сульфорафан және оның қатерлі ісікке, өлімге, қартаюға, ми мен мінез-құлыққа, жүрек ауруына және басқаларына әсері
Изотиоцианаттар сіздің диетаңызға түсетін ең маңызды өсімдік қосылыстарының бірі болып табылады. Бұл бейнеде мен олар үшін жасаған ең жан-жақты жағдайды жасаймын. Қысқаша назар аударыңыз ба? Төмендегі уақыт нүктелерінің бірін таңдап, сүйікті тақырыпқа өтіңіз. Толық уақыт шкаласы.
Негізгі бөлімдер:
00: 01: 14 - қатерлі ісік және өлім
00: 19: 04 - қартаю
00: 26: 30 - Ми және мінез
00: 38: 06 - Соңғы қорытынды
00: 40: 27 - Доза
Толық уақыт шкаласы:
00: 00: 34 - бейнефильмнің басты тақырыбы - сульффоран енгізу.
00: 01: 14 - Өсімдік тұтыну және барлық өлім-жітімде қысқарту.
00: 02: 12 - простата қатерлі ісігі.
00: 02: 23 - Мочевина рагы қатер.
00: 02: 34 - темекі шегудегі қатерлі ісік.
00: 02: 48 - Сүт безі қатерлігі қатері.
00: 03: 13 - Гипотетикалық: егер сізде рак бар болса? (интервенционалдық)
00: 03: 35 - қатерлі ісік пен өлім қауымдасқан деректерді басқаратын шынайы механизм.
00: 04: 38 - сульфорфан және қатерлі ісік.
00: 05: 32 - егеуқұйрықтардағы ісік ісігін дамытуға арналған брокколи өсімдік сығындысының күшті әсерін көрсететін жануарлы дәлел.
00: 26: 30 - мидың және мінез-құлық бөлімінің басы.
00: 27: 18 - аутизмге арналған брокколи өсімдік сығындысының әсері.
00: 27: 48 - глюкурофенияларға арналған глюкурофанның әсері.
00: 28: 17 - Депрессияның талқылануын бастау (ықтимал механизм мен зерттеулер).
00: 31: 21 - Стресс-индуцирленген депрессияның 10 түрлі модельдерін қолданып тышқандарды зерттеу, сульфарфетан сияқты флоксин (prozac) сияқты тиімді.
00: 32: 00 - зерттеу тышқандарда глюкарофаниннің тікелей ішуін көрсетеді, социологиялық жеңілдетудің стресс-моделінен депрессияны болдырмау үшін де тиімді.
00: 33: 01 - нейродегенерация бөлімінің басы.
00: 33: 30 - сульфорфан және Альцгеймер ауруы.
00: 33: 44 - Сульфорфан және Паркинсон ауруы.
00: 33: 51 - Сульфорфан және Хантингтон ауруы.
00: 34: 13 - Сульфорфан жылу әсерін арттырады.
00: 34: 43 - бас миының зақымдану секциясының басы.
00: 35: 01 - Туберкулезге қарсы тестілеуден кейін бірден сульфорфан препараты (тышқанның зерттеуі).
00: 35: 55 - сульфорфан және нейрондық пластика.
00: 36: 32 - Сулфорафан тышқандар типіндегі II типті қант диабеті моделін үйренуді жақсартады.
00: 37: 19 - сульфорфан және дученне бұлшықет дистрофиясы.
00: 38: 06 - Кешіктіріп бейнелеу: өлім және қатерлі ісік, ДНҚ зақымдануы, тотығу стресті және қабыну, бензолдың экскрециясы, жүрек-қан тамырлары аурулары, II типті диабет, миға әсер ету (депрессия, аутизм, шизофрения, нейрогенерация), NRF2 жолдары.
00: 40: 27 - брокколи өскіндер немесе сульффоранның дозасын анықтау туралы ойлар.
00: 41: 01 - Үйде өсіруге арналған анекдоттар.
00: 43: 14 - Пісіру температуралары және сульфарфан белсенділігі туралы.
00: 43: 45 - Глюкорафаниннен сульффоранның Gut бактерияларына айналуы.
00: 44: 24 - Көкөністерден белсенді мирозиназмен үйлескен кезде қоспалар жақсы жұмыс істейді.
00: 44: 56 - пісіру әдісі және крест тәрізді көкөністер.
2 (NF-E2) байланысқан эритроиды-ядролық факторы 2 факторы болып табылады, әйтпесе Nrf2 ретінде белгілі, антиоксиданттық және детоксикациялы ферменттердің түрлерін реттейтін транскрипция факторы. Зерттеулерде тотығу стресстерін бақылаудағы рөлі де көрсетілген. Альцгеймер ауруы және Паркинсон ауруы сияқты нейродегенеративті аурулардың көпшілігі тотығу стресімен және созылмалы қабынуымен сипатталады, жалпы мақсаттары Nrf2 емдеу тәсілдері. Доктор Алекс Хименес DC, CCST Insight
Қорытынды ескертулер
Транскрипция коэффициенті NRF2 UPR, UPS және autophagy өзгерістерін сезіну және модуляциялау арқылы протеостатикалық жауапты орындайды (Cурет 4). Нәтижесінде, NRF2 болмауы белоктың потенциалын күшейтеді, бұл NRF2 оңтайлы ақуызды тазарту үшін қажет екенін көрсетеді. Барлық бірге NRF2 протеинопатиялар үшін қызықты терапиялық мақсат болуы мүмкін екендігін болжауға болады.
Сурет 4 NRF2 - протеотоксичтен алынған шұғыл сигналдарды қорғаныстық транскрипциялық жауапқа байланыстыратын хаб ретінде. Ашылған / қатпарланбаған ақуыздардың жиналуы ER-дағы ақуыздың реакциясының (UPR) активтендіруіне әкеледі. PERK немесе MAPK белсендіру ER-резиденті Gpx8 және GSH деңгейлерін реттейтін бірнеше ферменттердің транскрипционды индукциясын әкелуі мүмкін, бұл дұрыс протеинді қалыптастыруды қамтамасыз ету үшін маңызды. Протеиннің агрегаттары протеазомдық белсенділікті (UPS) тежейді, NRF2 деградациясынан аулақ болар. NRF2-те Psma3, Psma6, Psmb1, Psmb5 және Pomp гендерінің транскрипциясын арнайы түрде көрсету үшін көрсетілген. Бірнеше басқа бөлімшелер NRF2-ға тәуелді түрде D3T-ке жауап ретінде реттеліп, NRF2-мен реттелетін протеасомдық субөңірлердің тізімін кеңейтуі мүмкін. Автофазия ақуыздың агрегаттарының тозуының негізгі жолдары болып табылады. Autophagy сондай-ақ NRF2-ты реттейді, NRF2 транскрипционды индукциямен p62, Ndp52, Ulk1, Atg2b, Atg4c, Atg5, Atg7 және Gabarapl1 арқылы осы бұзылу жолын жалғайды.
Жоғарыда келтірілген мақалаға сәйкес, невродегенеративті аурулардың белгілері әртүрлі емдеу әдістерімен емделе алса, зерттеу жұмыстары Nrf2 активтендіру емдеудің пайдалы тәсілі бола алатындығын көрсетті. Өйткені Nrf2 белсендірушілері аурудың кең тетіктерін көздейді, барлық нейродегенеративті аурулар Nrf2 транскрипция факторын пайдаланудан пайда көре алады. Nrf2 нәтижелері нейродегенеративті ауруларды емдеуде төңкеріс жасады. Біздің ақпаратымыздың көлемі хиропротикалық және омыртқаның денсаулығына қатысты мәселелермен шектеледі. Тақырыпты талқылау үшін доктор Хименеске хабарласыңыз немесе бізге телефон арқылы хабарласыңыз915-850-0900 .
Қосымша тақырыпты талқылау: Тікелей ауырсынуды хирургсыздандыру
Тізедегі ауырсыну - бұл белгілі симптом, ол әртүрлі тізе жарақаттарына және / немесе жағдайларға байланысты болуы мүмкін, соның ішіндеспорттық жарақаттар. Тізе - адам ағзасындағы ең күрделі буындардың бірі, өйткені ол төрт сүйектің, төрт байламның, әртүрлі сіңірлердің, екі менисканың және шеміршектің қиылысуынан тұрады. Американдық жанұялық дәрігерлер академиясының мәліметі бойынша, тізедегі ауырсынудың ең көп тараған себептеріне пателлярлық сублаксация, пателлярлық тендинит немесе секіргіш тізе және Осгуд-Шлаттер ауруы жатады. Тізедегі ауырсыну 60 жастан асқан адамдарда жиі кездесетініне қарамастан, тізе ауруы балалар мен жасөспірімдерде де болуы мүмкін. Тізедегі ауырсынуды RICE әдістері бойынша үйде емдеуге болады, дегенмен ауыр тізе жарақаттары шұғыл медициналық көмекті, соның ішінде хиропротикалық күтімді қажет етуі мүмкін. �
Тотығу стрессі бос радикалдар немесе тұрақсыз молекулалар тудыратын жасуша зақымдануы ретінде сипатталады, бұл сайып келгенде сау функцияға әсер етуі мүмкін. Адам ағзасы бактериялар мен вирустарды бейтараптандыру үшін бос радикалдарды жасайды, бірақ оттегі, ластану және радиация сияқты сыртқы факторлар жиі бос радикалдарды тудыруы мүмкін. Тотығу стрессі көптеген денсаулық мәселелерімен байланысты болды.
Тотығу стресстері және басқа стресс факторлары адам ағзасының антиоксиданттық реакциясын реттеуге көмектесетін ішкі қорғаныс механизмдерін қосады. Nrf2 - тотығу стресінің деңгейін сезетін және жасушаларға ішкі және сыртқы факторлардан өзін қорғауға мүмкіндік беретін ақуыз. Nrf2 антиоксиданттық ферменттер мен стресс-жауап гендерін өндіруге қатысатын гендерді реттеуге көмектесетіні де көрсетілді. Төмендегі мақаланың мақсаты - түсіндіру Nrf2 әсерлері қатерлі ісік кезінде.
дерексіз
Keap1-Nrf2 жолы тотығу және электрофильдік стресске цитопротекторлық жауаптардың негізгі реттеушісі болып табылады. Nrf2 транскрипция факторы арқылы іске қосылған жасушалық сигналдық жолдар қалыпты және қатерлі тіндерде қатерлі ісіктің басталуына және прогрессіне жол бермесе де, толық қатерлі жасушаларда Nrf2 белсенділігі қатерлі ісікке төзімділікті арттыру және ісік жасушаларының өсуін арттыру арқылы өсу артықшылығын қамтамасыз етеді. Бұл графикалық шолуда біз Keap1-Nrf2 жолына және оның рак клеткаларындағы реттелуінің бұзылуына шолу жасаймыз. Біз сондай-ақ рак клеткаларындағы конститутивтік Nrf2 белсендіруінің салдарын және оны рак гендік терапиясында қалай пайдалануға болатынын қысқаша қорытындылаймыз.
Түйінді сөздер:Nrf2, Keap1, қатерлі ісік, антиоксиданттық жауап элементі, гендік терапия
кіріспе
Keap1-Nrf2 жолы реактивті оттегі түрлері (ROS) және электрофилдер [1] тудыратын эндогендік және экзогендік кернеулерге цитопротекторлық жауаптардың негізгі реттеушісі болып табылады. Жолдағы негізгі сигналдық белоктар мақсатты гендердің реттеуші аймақтарындағы антиоксиданттық жауап элементімен (ARE) шағын Maf ақуыздарымен байланысатын Nrf2 (ядролық фактор эритроидты 2-байланысты фактор 2) және Keap1 (Kelch ECH) болып табылады. байланыстырушы ақуыз 1), Nrf2-мен байланысатын және оның убиквитин протеазома жолы арқылы ыдырауына ықпал ететін репрессорлық ақуыз (1-сурет). Keap1 - цистеинге өте бай ақуыз, Keap1 тінтуірінде барлығы 25 және адамда 27 цистеин қалдығы бар, олардың көпшілігін in vitro жағдайында әртүрлі тотықтырғыштар мен электрофилдер арқылы өзгертуге болады [2]. Осы қалдықтардың үшеуі, C151, C273 және C288, Nrf1 ядролық транслокациясына және кейінгі мақсатты ген экспрессиясына әкелетін Keap2 конформациясын өзгерту арқылы функционалды рөл атқаратыны көрсетілген [3] (1-сурет). Keap1-дегі цистеин модификацияларының Nrf2 активациясына әкелетін нақты механизмі белгісіз, бірақ екі басым, бірақ бір-бірін жоққа шығармайтын модель (1) Keap1 IVR-де орналасқан тиол қалдықтарындағы Keap1 модификациялары болатын «ілмек және ілмек» моделі болып табылады. Nrf2-мен өзара әрекеттесуін бұзады, бұл Nrf2 ішінде полиубиквитинизацияланбайтын лизин қалдықтарының сәйкессіздігін тудырады және (2) тиол модификациясы Cul3-тің Keap1-ден диссоциациялануын тудыратын модель [3]. Екі модельде де индуктормен модификацияланған және Nrf2-байланысқан Keap1 инактивтендірілген және, демек, жаңадан синтезделген Nrf2 ақуыздары Keap1-ді айналып өтіп, ядроға ауысады, ARE-мен байланысады және NAD(P)H сияқты Nrf2 мақсатты гендерінің экспрессиясын басқарады. хиноноксидоредуктаза 1 (NQO1), гемоксигеназа 1 (HMOX1), глутамат-цистеин лигаза (GCL) және глутатион S трансферазалары (GSTs) (2-сурет). Nrf1 мақсатты генінің индукциясына әкелетін Keap2 тиолдарының модификацияларына қосымша, p21 және p62 сияқты ақуыздар Nrf2 немесе Keap1-ге қосыла алады, осылайша Nrf2 және Keap1 [1], [3] (3-сурет) өзара әрекеттесуін бұзады.
1-сурет. Nrf2 және Keap1 құрылымдары және цистеин коды. (A) Nrf2 589 аминқышқылдарынан тұрады және алты эволюциялық жоғары сақталған Neh1-6 домені бар. Neh1 құрамында bZip мотиві, негізгі аймақ – лейцин сыдырма (L-Zip) құрылымы бар, мұнда негізгі аймақ ДНҚ тану үшін жауап береді және L-Zip шағын Maf ақуыздарымен димеризацияға делдал болады. Neh6 ядродағы Nrf2 деградациясына делдалдық дегрон ретінде қызмет етеді. Neh4 және 5 - транзакция домендері. Neh2 құрамында Keap1-пен әрекеттесу үшін қажет ETGE және DLG мотивтері және Nrf7-нің Keap1-тәуелді полиубиквитинациясы мен ыдырауы үшін қажетті лизин қалдықтарының гидрофильді аймағы (2 K) бар. (B) Keap1 624 амин қышқылы қалдықтарынан тұрады және бес доменге ие. Протеин мен ақуыздың өзара әрекеттесуінің екі мотиві, BTB домені және Келч домені аралық аймақпен (IVR) бөлінген. BTB домені IVR N-терминалды бөлігімен бірге Keap1 гомодимерленуіне және Куллин3 (Cul3) байланысуына делдал болады. Kelch домені және C-терминал аймағы Neh2-мен өзара әрекеттесуде делдалдық етеді. (C) Nrf2 Neh1 ETGE және DLG мотивтері арқылы Keap2 екі молекуласымен әрекеттеседі. ETGE және DLG екеуі де Keap1 Kelch мотивінің төменгі бетіндегі ұқсас тораптармен байланысады. (D) Keap1 цистеин қалдықтарына бай, адам ақуызында 27 цистеин бар. Бұл цистеиндердің кейбірі негізгі қалдықтардың жанында орналасқан, сондықтан электрофилдер мен тотықтырғыштардың тамаша нысанасы болып табылады. Цистеин қалдықтарының электрофилдер арқылы модификациясының үлгісі цистеин коды деп аталады. Цистеин коды гипотезасы құрылымдық жағынан әртүрлі Nrf2 белсендіруші агенттердің әртүрлі Keap1 цистеиндеріне әсер ететінін ұсынады. Цистеин модификациялары Nrf1 DLG және Keap2 Kelch домендері арасындағы өзара әрекеттесуді бұзатын Keap1 конформациялық өзгерістеріне әкеледі, осылайша Nrf2 полиубиквитинациясын тежейді. Cys151, Cys273 және Cys288 функционалдық маңыздылығы көрсетілді, өйткені Cys273 және Cys288 индукторлармен Nrf2 белсендіруі үшін Nrf151 және Cys2 басу үшін қажет [1], [3].
2-сурет. Nrf2-Keap1 сигналдық жолы. (A және B) базальды жағдайда екі Keap1 молекуласы Nrf2-мен байланысады және Nrf2 Cul3 негізіндегі E3 лигаза кешенімен полиубиктилденеді. Бұл полиубиквилизация протеазома арқылы Nrf2 жылдам деградациясына әкеледі. Nrf2 аз бөлігі ингибиторлық кешеннен шығып кетеді және базальді ARE-тәуелді геннің экспрессиясына делдалдық ету үшін ядрода жиналады, осылайша жасушалық гомеостазды сақтайды. (C) Стресс жағдайында индукторлар ингибиторлық кешеннің диссоциациялануы арқылы Nrf1 убиквитиляциясының тежелуіне әкелетін Keap2 цистеиндерін өзгертеді. (D) Топса мен ысырма үлгісіне сәйкес Keap1 цистеинінің арнайы қалдықтарының модификациясы Keap1-де конформациялық өзгерістерге әкеледі, нәтижесінде Nrf2 DLG мотиві Keap1-ден ажырайды. Nrf2 убиквитинациясы бұзылған, бірақ ETGE мотивімен байланыс сақталады. (E) Keap1-Cul3 диссоциациясының моделінде Keap1 және Cul3 байланысуы электрофилдерге жауап ретінде бұзылады, бұл Nrf2-нің убиквитинация жүйесінен шығуына әкеледі. Ұсынылған модельдердің екеуінде де индуктормен модификацияланған және Nrf2-байланысқан Keap1 инактивацияланған және, тиісінше, жаңадан синтезделген Nrf2 ақуыздары Keap1-ді айналып өтіп, ядроға ауысады, антиоксиданттық әсер ету элементімен (ARE) байланысады және Nrf2 нысанасының экспрессиясын басқарады. NQO1, HMOX1, GCL және GSTs [1], [3] сияқты гендер.
Сурет 3. Қатерлі ісік кезінде Nrf2 конститутивтік ядролық жинақталу механизмдері. (A) Nrf2 немесе Keap1-дегі соматикалық мутациялар осы екі ақуыздың өзара әрекеттесуін бұзады. Nrf2-де мутациялар ETGE және DLG мотивтеріне әсер етеді, бірақ Keap1-де мутациялар біркелкі таралады. Сонымен қатар, KrasG12D[5] сияқты онкогенді белсендіру немесе PTEN [11] сияқты ісік супрессорларының бұзылуы Nrf2 транскрипциялық индукциясына және Nrf2 ядросының жоғарылауына әкелуі мүмкін. (B) Өкпе мен простата обырындағы Keap1 промоторының гиперметилденуі Keap1 мРНҚ экспрессиясының төмендеуіне әкеледі, бұл Nrf2 ядролық жинақталуын арттырады [6], [7]. (C) Отбасылық папиллярлы бүйрек карциномасында фумаратгидратаза ферментінің белсенділігінің жоғалуы фумараттың жинақталуына және одан әрі Keap1 цистеин қалдықтарының сукцинациясына әкеледі (2SC). Бұл трансляциядан кейінгі модификация Keap1-Nrf2 өзара әрекеттесуінің бұзылуына және Nrf2 ядролық жинақталуына әкеледі [8], [9]. (D) p62 және p21 сияқты бұзушы ақуыздардың жинақталуы Nrf2-Keap1 байланысуын бұзуы мүмкін және Nrf2 ядросының жоғарылауына әкеледі. p62 Nrf1 үшін байланыстыратын қалтамен қабаттасатын Keap2-мен байланысады және p21 Nrf2-нің DLG және ETGE мотивтерімен тікелей әрекеттеседі, осылайша Keap1-мен бәсекелеседі [10].
Қатерлі ісік ауруындағы Nrf2 белсендіру және реттелу механизмдері
Nrf2 белсендіру арқылы қамтамасыз етілген цитопротекция қалыпты және қатерлі тіндерде қатерлі ісік химиопрофилактикасы үшін маңызды болғанымен, толық қатерлі жасушаларда Nrf2 белсенділігі қатерлі ісікке төзімділікті арттыру және ісік жасушаларының өсуін арттыру арқылы өсу артықшылығын қамтамасыз етеді [4]. Nrf2 сигнал беру жолы әртүрлі қатерлі ісіктерде конститутивтік белсендірілетін бірнеше механизмдер сипатталған: (1) Keap1-дегі соматикалық мутациялар немесе Nrf1-нің Keap2 байланыстыру доменінде олардың өзара әрекеттесуін бұзады; (2) Nrf1-нің ақаулы репрессиясына әкелетін Keap2 экспрессиясының эпигенетикалық үнсіздігі; (3) Keap62-Nrf1 кешенінің диссоциациясына әкелетін p2 сияқты бұзушы ақуыздардың жинақталуы; (4) онкогенді K-Ras, B-Raf және c-Myc арқылы Nrf2 транскрипциялық индукциясы; және (5) фумаратгидратаза ферментінің белсенділігін жоғалтуға байланысты отбасылық папиллярлы бүйрек карциномасында пайда болатын сукцинилдену арқылы Keap1 цистеиндерінің трансляциядан кейінгі модификациясы [3], [4], [5], [6], [7], [ 8], [9], [10] (Cурет 3). Құрамында мол Nrf2 протеині дәрілік зат алмасуға қатысатын гендердің экспрессиясының жоғарылауын тудырады, осылайша химиотерапиялық препараттарға және сәулелік терапияға төзімділікті арттырады. Сонымен қатар, жоғары Nrf2 ақуыз деңгейі қатерлі ісік ауруының нашар болжамымен байланысты [4]. Шамадан тыс белсенді Nrf2 сонымен қатар глюкоза мен глутаминді пурин синтезін күшейтетін анаболикалық жолдарға бағыттау және жасуша пролиферациясын ынталандыру үшін пентозофосфат жолына әсер ету арқылы жасуша пролиферациясына әсер етеді [11] (4-сурет).
4-сурет. Ісік пайда болудағы Nrf2 қос рөлі. Физиологиялық жағдайларда жасушалық гомеостазды сақтау үшін Nrf2 ядросының төмен деңгейі жеткілікті. Nrf2 канцерогендерді, ROS және басқа ДНҚ-зақымдаушы агенттерді жою арқылы ісіктің басталуын және ісік метастазын тежейді. Ісік пайда болу кезінде жинақталған ДНҚ зақымдануы Nrf2 конститутивтік гиперактивтілігіне әкеледі, бұл автономды қатерлі жасушаларға эндогендік ROS жоғары деңгейіне төтеп беруге және апоптозды болдырмауға көмектеседі. Тұрақты жоғарылаған ядролық Nrf2 деңгейлері метаболизмді қайта бағдарламалауға және жасуша пролиферациясын жақсартуға ықпал ететін цитопротекторлық гендерге қосымша метаболикалық гендерді белсендіреді. Nrf2 деңгейі жоғары қатерлі ісіктер радио және химорезистенттілік пен агрессивті рак клеткаларының көбеюіне байланысты нашар болжаммен байланысты. Осылайша, Nrf2 жолының белсенділігі ісік пайда болуының бастапқы кезеңдерінде қорғаныш болып табылады, бірақ кейінгі кезеңдерінде зиянды. Сондықтан қатерлі ісіктің алдын алу үшін Nrf2 белсенділігін арттыру маңызды тәсіл болып қала береді, ал қатерлі ісік ауруын емдеу үшін Nrf2 тежелуі қажет [4], [11].
Жоғары Nrf2 белсенділігі әдетте қолайсыз нәтижелері бар рак жасушаларында болатынын ескере отырып, Nrf2-ні тежейтін терапия қажет. Өкінішке орай, кейбір басқа bZip отбасы мүшелерімен құрылымдық ұқсастығына байланысты, арнайы Nrf2 ингибиторларын әзірлеу қиын міндет болып табылады және Nrf2 тежелуіне қатысты бірнеше зерттеулер ғана жарияланған. Табиғи өнімдерді скрининг арқылы Рен және т.б. [12] цисплатиннің химиотерапиялық тиімділігін арттыратын Nrf2 ингибиторы ретінде ісікке қарсы брусатол қосылысын анықтады. Сонымен қатар, PI3K тежегіштері [11], [13] және Nrf2 siRNA [14] ісік жасушаларында Nrf2 тежеу үшін қолданылған. Жақында біз Nrf2 деңгейі жоғары қатерлі ісік жасушаларын нысанаға алу үшін қатерлі ісік суицидінің гендік терапиясы деп аталатын балама тәсілді қолдандық. Құрамында тимидинкиназа (TK) бар Nrf2 басқаратын лентивирустық векторлар [15] жоғары ARE белсенділігі бар рак жасушаларына тасымалданады және жасушалар пропрепаратпен, Ганцикловирмен (GCV) өңделеді. GCV GCV-монофосфатқа метаболизденеді, ол одан әрі жасушалық киназалармен улы трифосфатты түрге дейін фосфорланады [16] (5-сурет). Бұл ісік жасушалары бар TK ғана емес, сонымен қатар бақылаушы әсерге байланысты көрші жасушаларды тиімді өлтіруге әкеледі [17]. ARE реттелетін TK/GCV гендік терапиясы қатерлі ісікке қарсы химиотерапевтік агент доксорубицинді емдеуге біріктіру арқылы одан әрі жақсартылуы мүмкін [16], бұл тәсіл дәстүрлі терапиямен бірге пайдалы болуы мүмкін деген түсінікті қолдайды.
5-сурет. Суицидтік гендік терапия. Қатерлі ісік жасушаларындағы Nrf2 конститутивтік ядролық жинақталуын Nrf2 басқаратын вирустық векторды қатерлі ісік суицидінің гендік терапиясы үшін пайдалану арқылы пайдалануға болады [16]. Бұл тәсілде төрт ARE бар минималды SV40 промоторы астында тимидинкиназаны (ТК) экспрессиялайтын лентивирустық вектор (LV) өкпе аденокарциномасының жасушаларына ауыстырылады. Nrf2 жоғары ядролық деңгейлері Nrf2 байланысуы арқылы TK сенімді экспрессиясына әкеледі. Содан кейін жасушалар пропрепаратпен өңделеді, TK арқылы фосфорланатын Ганцикловир (GCV). Трифосфорирленген GCV ДНҚ синтезін бұзады және бақылаушы әсерге байланысты құрамында ісік жасушалары бар TK ғана емес, сонымен қатар көрші жасушаларды да тиімді өлтіруге әкеледі.
Nrf2 - адам ағзасында тотығу стрессін жоюға көмектесетін күшті антиоксиданттардың өндірісін қоздыратын негізгі реттеуші. Nrf2 жолы арқылы супероксид дисмутаза немесе SOD, глутатион және каталаза сияқты әртүрлі антиоксиданттық ферменттер де белсендіріледі. Сонымен қатар, куркума, ашваганда, бакопа, жасыл шай және сүт ошаған сияқты кейбір фитохимиялық заттар Nrf2 белсендіреді. Зерттеулер мұны анықтады Nrf2 белсендіру табиғи түрде жасушалық қорғанысты күшейтіп, адам ағзасының тепе-теңдігін қалпына келтіре алады.
Доктор Алекс Хименес DC, CCST Insight
Сульфорафан және оның қатерлі ісікке, өлімге, қартаюға, ми мен мінез-құлыққа, жүрек ауруына және басқаларына әсері
Изотиоцианаттар сіздің диетаңызға түсетін ең маңызды өсімдік қосылыстарының бірі болып табылады. Бұл бейнеде мен олар үшін жасаған ең жан-жақты жағдайды жасаймын. Қысқаша назар аударыңыз ба? Төмендегі уақыт нүктелерінің бірін таңдап, сүйікті тақырыпқа өтіңіз. Толық уақыт шкаласы.
Негізгі бөлімдер:
00: 01: 14 - қатерлі ісік және өлім
00: 19: 04 - қартаю
00: 26: 30 - Ми және мінез
00: 38: 06 - Соңғы қорытынды
00: 40: 27 - Доза
Толық уақыт шкаласы:
00: 00: 34 - бейнефильмнің басты тақырыбы - сульффоран енгізу.
00: 01: 14 - Өсімдік тұтыну және барлық өлім-жітімде қысқарту.
00: 02: 12 - простата қатерлі ісігі.
00: 02: 23 - Мочевина рагы қатер.
00: 02: 34 - темекі шегудегі қатерлі ісік.
00: 02: 48 - Сүт безі қатерлігі қатері.
00: 03: 13 - Гипотетикалық: егер сізде рак бар болса? (интервенционалдық)
00: 03: 35 - қатерлі ісік пен өлім қауымдасқан деректерді басқаратын шынайы механизм.
00: 04: 38 - сульфорфан және қатерлі ісік.
00: 05: 32 - егеуқұйрықтардағы ісік ісігін дамытуға арналған брокколи өсімдік сығындысының күшті әсерін көрсететін жануарлы дәлел.
00: 26: 30 - мидың және мінез-құлық бөлімінің басы.
00: 27: 18 - аутизмге арналған брокколи өсімдік сығындысының әсері.
00: 27: 48 - глюкурофенияларға арналған глюкурофанның әсері.
00: 28: 17 - Депрессияның талқылануын бастау (ықтимал механизм мен зерттеулер).
00: 31: 21 - Стресс-индуцирленген депрессияның 10 түрлі модельдерін қолданып тышқандарды зерттеу, сульфарфетан сияқты флоксин (prozac) сияқты тиімді.
00: 32: 00 - зерттеу тышқандарда глюкарофаниннің тікелей ішуін көрсетеді, социологиялық жеңілдетудің стресс-моделінен депрессияны болдырмау үшін де тиімді.
00: 33: 01 - нейродегенерация бөлімінің басы.
00: 33: 30 - сульфорфан және Альцгеймер ауруы.
00: 33: 44 - Сульфорфан және Паркинсон ауруы.
00: 33: 51 - Сульфорфан және Хантингтон ауруы.
00: 34: 13 - Сульфорфан жылу әсерін арттырады.
00: 34: 43 - бас миының зақымдану секциясының басы.
00: 35: 01 - Туберкулезге қарсы тестілеуден кейін бірден сульфорфан препараты (тышқанның зерттеуі).
00: 35: 55 - сульфорфан және нейрондық пластика.
00: 36: 32 - Сулфорафан тышқандар типіндегі II типті қант диабеті моделін үйренуді жақсартады.
00: 37: 19 - сульфорфан және дученне бұлшықет дистрофиясы.
00: 38: 06 - Кешіктіріп бейнелеу: өлім және қатерлі ісік, ДНҚ зақымдануы, тотығу стресті және қабыну, бензолдың экскрециясы, жүрек-қан тамырлары аурулары, II типті диабет, миға әсер ету (депрессия, аутизм, шизофрения, нейрогенерация), NRF2 жолдары.
00: 40: 27 - брокколи өскіндер немесе сульффоранның дозасын анықтау туралы ойлар.
00: 41: 01 - Үйде өсіруге арналған анекдоттар.
00: 43: 14 - Пісіру температуралары және сульфарфан белсенділігі туралы.
00: 43: 45 - Глюкорафаниннен сульффоранның Gut бактерияларына айналуы.
00: 44: 24 - Көкөністерден белсенді мирозиназмен үйлескен кезде қоспалар жақсы жұмыс істейді.
00: 44: 56 - пісіру әдісі және крест тәрізді көкөністер.
Бұл жұмысқа Финляндия академиясы, Сигрид Юселиус қоры және Финляндияның онкологиялық ұйымдары қолдау көрсетті.
Қорытындылай келе, NFE2L2 немесе Nrf2 ретінде белгілі ядролық фактор (эритроидты 2) тәрізді 2 адам ағзасын тотығу стрессінен қорғайтын антиоксиданттардың өндірісін арттыратын ақуыз болып табылады. Жоғарыда сипатталғандай, Nrf2 жолын ынталандыру тотығу стрессінен туындаған ауруларды, соның ішінде қатерлі ісіктерді емдеуге арналған зерттеулер жүргізілуде. Біздің ақпаратымыздың көлемі хиропротикалық және омыртқаның денсаулығына қатысты мәселелермен шектеледі. Тақырыпты талқылау үшін доктор Хименеске хабарласыңыз немесе бізге телефон арқылы хабарласыңыз915-850-0900 .
Қосымша тақырыпты талқылау: Тікелей ауырсынуды хирургсыздандыру
Тізедегі ауырсыну - бұл белгілі симптом, ол әртүрлі тізе жарақаттарына және / немесе жағдайларға байланысты болуы мүмкін, соның ішіндеспорттық жарақаттар. Тізе адамның денесінде ең күрделі түйістердің бірі болып табылады, себебі ол төрт сүйек, төрт линза, әртүрлі сіңірлер, екі менисци және шеміршектің қиылысынан тұрады. Америка отбасылық дәрігерлер академиясының мәліметі бойынша, тізе ауруларының ең көп кездесетін себептері: пателярлық сублухация, пателлярлық тенденит немесе буынның тізесі және Осгуд-Шлаттер ауруы. Тізе ауруы, ең алдымен, 60 жастан асқан адамдарда кездессе де, балалар мен жасөспірімдерде тізе ауыруы мүмкін. Тізе ауруы үй жағдайында RICE әдістерімен емделе алады, бірақ ауыр тізе жарақаттары шұғыл көмекке, соның ішінде шипопротикалық күтімді қажет етеді.
ДНҚ шамамен 20,000 XNUMX генді қолдайды, олардың әрқайсысында салауатты өмір салты үшін қажетті ақуыз немесе фермент жасау бағдарламасы бар. Осы үлгілердің әрқайсысы әр заттың және/немесе химиялық заттардың қанша мөлшері түзілетінін және олардың қандай жағдайда дамитынын басқаратын белгілі бір «промотор» арқылы үнемі реттелуі керек.
Антиоксидантты әсер ету элементі немесе ARE деп аталатын қосқыш тәрізді промотор аймақтарының белгілі бір түріне қосылу арқылы Nrf2 факторыЖасушалардың стресстік жағдайларда өмір сүруіне мүмкіндік беретін жүздеген ерекше гендердің жасалу жылдамдығын қолдайды. Содан кейін бұл гендер тотықтырғыштарды бейтараптандыру және олардың өндірісінде қалдырылған улы жанама өнімдерді тазарту арқылы қорғаныс желісін дамытатын антиоксиданттық ферменттердің таңдауын жасайды, сонымен қатар олар келтірген зақымдарды қалпына келтіруге көмектеседі.
Тотығу стрессі дегеніміз не?
Супероксид радикалы немесе O2- және сутегі асқын тотығы немесе H2O2 сияқты бірнеше тотықтырғыштар адам ағзасын қолдайтын заттарды және/немесе химиялық заттарды жағу тәжірибесі арқылы жасалған. Адам ағзасында біз тұтынатын реактивті тағамдар мен сусындарды бейтараптандыратын және детоксикациялайтын антиоксиданттық ферменттер бар. Nrf2 тепе-теңдікті сақтау үшін олардың өндірісін модуляциялайды және барлық осы ферменттерге сұранысты баса көрсетеді. Бұл тепе-теңдік бірнеше факторлармен, соның ішінде жаспен үзілуі мүмкін.
Қартайған сайын адам ағзасы Nrf2-ні азырақ жасайды және бұл нәзік тепе-теңдік бірте-бірте тотығу жағына қарай бұрыла бастайды, бұл күй тотығу стрессі деп аталады. Ауру сонымен қатар тотықтырғыштардың артық өндірілуіне себеп болуы мүмкін. Инфекциялар, аллергиялар және аутоиммундық бұзылулар біздің иммундық жасушаларды О2- сияқты реактивті тотықтырғыштарды жасау үшін қосымша қоздыруы мүмкін. , H2O2, OH және HOCl, мұнда сау жасушалар зақымдалады және қабынумен жауап береді. Қартаюмен байланысты аурулар, соның ішінде инфаркт, инсульт, қатерлі ісік және Альцгеймер ауруы сияқты нейродегенеративті жағдайлар да стрессті және қабыну реакциясын тудыратын тотықтырғыштардың дамуын арттырады.
Nrf2 активаторлары дегеніміз не?
Nrf2 протеині ферменттер мен гендерді қолдап, басқара алатындығына байланысты транскрипция факторы деп те аталады, когнитивті тепе-теңдік пен тотығу тепе-теңдігінің модификацияларына жауап беретін жасуша ішіндегі биохимиялық реакциялар тізбегінің құпия элементі болып табылады. Бұл жолдың сезімтал элементтері Nrf2-ні өзгертеді және разрядтайды, бұл оны ДНҚ-ға қарай жасушаның ядросына таралу үшін іске қосады. Nrf2 балама түрде жасушаны қорғау үшін қолдайтын гендер мен ферменттерді қосуы немесе өшіруі мүмкін.
Бақытымызға орай, Nrf2 белсендіргіштері болып табылатын әртүрлі заттар бірнеше ғасырлар бұрын қытайлық және жергілікті американдық дәстүрлі емдеу құралдарында қолданылған белгілі бір өсімдіктер мен сығындыларды тұтыну арқылы дамиды. Бұл фитохимиялық заттар бүгінде қолданылып жүрген Nrf2-белсендендіруші фармацевтикалық өнімдер сияқты жанама әсерлері азырақ күшті болып көрінеді.
Ядролық фактор эритроид 2-байланысты фактор, әдетте Nrf2 ретінде белгілі, гендерді, ферменттерді және антиоксиданттық реакцияларды реттеу арқылы жасушаны қорғайтын транскрипция факторы. Транскрипция факторлары - белгілі бір заттар мен химиялық заттардың, соның ішінде глутатион S-трансферазаларының немесе GST-тің пайда болуына ықпал ету үшін ДНҚ-ға қосылатын ақуыз түрі. Nrf2 белсендіру тотығу стрессін азайтуға көмектесетін күшті антиоксиданттық қабілетті көрсететін белсенді ақуыздардың өндірісін тудырады.
Доктор Алекс Хименес DC, CCST Insight
Nrf2 белсендіруінің артындағы ғылым
2 жылы Nrf2004-белсендендірудің бастапқы тағамдық қоспасы жасалғаннан кейін, Nrf2 жолының функциясына қатысты ең аз ақпарат белгілі болды. Nrf200 әдебиетінде ядролық фактор тәрізді 2 немесе NFE2L2 деп аталатын 2-ге жуық газет бар және зерттеушілер сүтқоректілердегі Nrf2 антиоксиданттық реакциясын енді ғана аша бастады. Алайда 2017 жылғы жағдай бойынша осы «бас реттеуші» бойынша 9,300-ден астам академиялық зерттеулер басылып шықты.
Шындығында, Nrf2 гендермен байланысы жоқ көптеген антиоксиданттық ферменттерді реттейді, оның орнына олар сау және патологиялық жағдайларда жасушалар, органдар және, сайып келгенде, организмдер кездесетін стресске байланысты әртүрлі жағдайлардан қорғауды ұсынады. Жарияланған академиялық зерттеулерден алынған ақпараттың осы жаңа көлеміне сүйене отырып, зерттеушілер енді жақсырақ дами алады Nrf2 диеталық қоспалар.
2007 жылдан бастап зерттеулер Nrf2 жолының күрделі қызметін көрсетті. Nrf2 активаторлары адам ағзасындағы әртүрлі құрылымдардың факторларына еліктейтіні анықталды. Осы жолдар арқылы Nrf2 активаторлары тепе-теңдікті сақтау және гендердің дамып келе жатқан талаптарына жауап беру үшін бүкіл жасушадағы өзгермелі жағдайларды сезіну үшін жабдықталған.
Неліктен Nrf2 белсендіретін қоспаларды пайдалану керек?
Nrf2-белсендендіру қабілеттері ағзалардың жасына қарай азайған сайын, өзгерістер орын алуы мүмкін. Зерттеулер көрсеткендей, жасушалардағы Nrf2 фокусы жас ұлғайған сайын азаяды, бұл тотығу стрессінің жоғарылауы маркерлерін көрсетеді. Атеросклероз және жүрек-тамыр аурулары, артрит, қатерлі ісік, семіздік, 2 типті қант диабеті, гипертония, катаракта және Альцгеймер ауруы, сондай-ақ Паркинсон ауруы сияқты әртүрлі жасқа байланысты аурулар осы өзгерістерге байланысты дамуы мүмкін. Осы денсаулық мәселелерімен тотығу стрессі анықталды.
Nrf2 активаторларының өндірісін арттыру үшін жасушаның қабілетін ынталандыру арқылы, Nrf2 диеталық қоспалар адам ағзасының тотығу стресінің әсеріне қарсы тұру қабілетін қалпына келтіруге көмектесе алады. Полиқанықпаған май қышқылдары немесе PUFAs, ең оңай тотығатын молекулалардың бірі болып табылады және олар бос радикалдардың зақымдалуына өте осал. Тиобарбитур қышқылы немесе TBARS өндірісі жасына қарай ұлғаюы мүмкін, бұл Nrf2 реттелетін жолдардың төмендеуімен бірге тотығу стрессінің жоғарылауын көрсетеді.
Биологиялық тұрғыдан гендік индукция шынымен баяу механизм болып табылады, әдетте жол арқылы тасымалдау үшін сағаттар қажет. Нәтижесінде, көптеген ферменттерде әртүрлі реттеуші ферменттер бірнеше минут ішінде іске қосылуы мүмкін өздерінің қосу/өшіру қосқыштары болады. Зерттеушілер белсендірудің осы білім қорын пайдаланатын Nrf2 активаторларының меншікті композицияларын әзірледі. Nrf2 белсендіру Nrf2 транскрипция факторының ингибиторынан босатылып, жасуша ядросына көшуінен ғана емес, сонымен қатар цитопротекторлық геннің экспрессиясын ынталандыру үшін арнайы ДНҚ тізбегімен байланысудан, Nrf2 ядродан шығарылатын қарқынды реттейді.
Адам ағзасындағы Nrf2 жою процедурасын және белсендіруін түсіну зерттеушілерге оның модуляциясы арқылы гендердің шағылысуын жүзеге асыру үшін әртүрлі Nrf2 активаторларының комбинацияларын құруға мүмкіндік берді. Білім базасының үйлесімі және басқа да көптеген зерттеулердің жиынтығы тағамдық қоспалар ретінде пайдалану үшін Nrf2 активаторларын шығаруға көмектесті. Біздің ақпаратымыздың көлемі хиропротикалық және омыртқаның денсаулығына қатысты мәселелермен шектеледі. Тақырыпты талқылау үшін доктор Хименеске хабарласыңыз немесе бізге телефон арқылы хабарласыңыз915-850-0900 .
Доктор Алекс Хименес мырзаның жетекшілігімен
Қосымша тақырыпты талқылау: Тікелей ауырсынуды хирургсыздандыру
Тізедегі ауырсыну - бұл белгілі симптом, ол әртүрлі тізе жарақаттарына және / немесе жағдайларға байланысты болуы мүмкін, соның ішіндеспорттық жарақаттар. Тізе адамның денесінде ең күрделі түйістердің бірі болып табылады, себебі ол төрт сүйек, төрт линза, әртүрлі сіңірлер, екі менисци және шеміршектің қиылысынан тұрады. Америка отбасылық дәрігерлер академиясының мәліметі бойынша, тізе ауруларының ең көп кездесетін себептері: пателярлық сублухация, пателлярлық тенденит немесе буынның тізесі және Осгуд-Шлаттер ауруы. Тізе ауруы, ең алдымен, 60 жастан асқан адамдарда кездессе де, балалар мен жасөспірімдерде тізе ауыруы мүмкін. Тізе ауруы үй жағдайында RICE әдістерімен емделе алады, бірақ ауыр тізе жарақаттары шұғыл көмекке, соның ішінде шипопротикалық күтімді қажет етеді.
Антиоксиданттар адам ағзасындағы тотығу процесін шектейтін қосылыстар деп аталады, егер бақылаусыз қалса, ол жасушалық зақым келтіруі мүмкін көптеген тізбекті реакцияларды дамыта алатын еркін радикалдарды жасай алады. Бақытымызға орай, адам денесі осындай иммундық механизмдерді жасай алады, алайда, реактивті оттегі түрлерін немесе ROS-ны монтаждау кезінде бейтараптандырылмайды, оттегімен тұндырылған кезде бақылаудан шығатын күлгін жалын қарастырылады, .
Жалынның метафорасын кеңейтуді жалғастыру үшін, ROS әсерін бейтараптандыруға қабілетсіз түпкілікті өнімі немесе реактивтік оттегі түрлерінің зақымдалуы, сонымен қатар қабыну, яғни адам денесі өрт болып табылады. Фантастикалық нәрсе - бұл денсаулыққа қарсы күресте үлкен көмекке ие антиоксиданттар, ал бұл антиоксидант - глютатион. 1889-те табылғанымен, глутатионның антиоксиданттық әсері заманауи зерттеулердегі ең қызықты тақырыптардың бірі болды.
Антиоксиданттар шебері: Глутатион
Қуатты зат - бұл цистеин, глутамин қышқылы және глициннен түзілетін трипептид. Глутатион адам ағзасын бос радикалдардың пайда болуынан қорғауға қабілетті болғандықтан, сайып келгенде иммундық жүйені сауықтыруға көмектеседі. Негізделген 2015 ішіндегі ғылыми есептерглютатионның пероксиринмен және каталазамен синергетикалық түрде әрекет ету қабілетін сутегі асқынына қарсы жасушаларды қорғауға көмектесетіні анықталды. Бұл синергетикалық формула реактивті оттегі түрлеріне немесе ROS-ға қарсы әрекет етеді. Глютатион, пероксиддрин және каталаза клеткалық гомеостаздың өсуінде маңызды элементтер болып табылады, ол сау клеткалардың, тіндердің және органдардың толықтай маңызды процесі болып табылады.
Сонымен қатар, глутатион иммундық жүйенің құрылымын және функциясын лимфоциттердің функцияларына маңызды әсерін арттырады. Сәйкесінше Иммунохимия кафедрасы, адам ағзасындағы глутатион деңгейін дұрыс толықтыру иммундық реакцияларды айтарлықтай арттырады. Мысалы, екі рандомизацияланған плацебо-бақыланатын сынақтар N-ацетилцистеинмен немесе NAC-пен иммунды бұзылған науқастарды терапиялық емдеудің екі жағдайда да иммунологиялық процестердің көпшілігінде айтарлықтай өсуіне әкелді, табиғи киллерлік жасушалардың белсенділігі. N-ацетилцистеин немесе NAC күкіртті глютатионнан пайдаланады және оны суда еритін және адам ағзасына шығаратын улы молекулалармен біріктіреді.
Глутатион сондай-ақ липок қышқылын жандандырады, сондай-ақ еркін радикалдарды бейтараптандыру үшін электронды жіберу арқылы белгілі бір жүйелік процестерді бастау үшін қажетті В және С дәрумендерін қайта өңдеу мүмкіндігіне ие. Зерттеу жұмысының негізінде PLOS ONE, глутатионды диабет метилусымен немесе T2DM және микобактерия туберкулезімен ауыратын науқастар әсер етті. Әдетте әлсіз иммундық жүйелері бар адамдарға M. tb немесе mycobacterium tuberculosis, ауру немесе инфекцияның көп әсерін көрсету үрдісі бар. Сонымен қатар, Type 2 қант диабеті метиллюл немесе T2DM бар адамдар T2DM жоқ адамдарға қарағанда екі-үш есе көп ТБ-ға бейім. Зерттеу жұмысы T2DM бар науқастардан оқшауланған макрофагтардың глутатион деңгейін жоғарылату M.Tb ауруы немесе инфекциясының бақылауын жақсартуға әкеліп соқтырды. Бұл нәтижелер T2DM бар науқастардағы глутатионның төменгі деңгейінің M. tb ауруы немесе инфекциясының жоғарылауына ықпал ететіндігін көрсетеді. Сонымен қатар, тәуелді Бритон және Сассекс медициналық мектебінде Диетро Геззи, тотығу стрессі ақыр аяғында иммундық жүйенің нашар құрылымы мен функциясын тудыруы мүмкін.
Бақытымызға орай, глутатион иммунитетті нығайту мен бақылауда маңызды рөл атқарады. Мысалы, глутатион иммундық жүйедегі туберкулезді және адаптивті процестерге, оның ішінде Т-лимфоциттердің пролиферациясына, полиморфонуклеарлы нейтрофилдердің фагоцитарлық белсенділігіне және дендритикалық жасушалық функцияларға маңызды болып табылады, олар негізі болуы мүмкін, себебі олар антигенді ұсынатын жасушалардан тұрады . Cell-meditated иммунитеті бастапқыда макрофагтардың және дендрит клеткаларының эндокристалды весикулярларында нашарлауы басталып, антигендерге тән Т-клеткаларының пролиферациясын белсендіру үшін кішірек пептидтер көрсетіледі. Сонымен қатар, глутатион цитокиндердің пайда болуына көмектеседі және микробактерияны қоса алғанда, жасушааралық патогенділерден қорғау үшін маңызды болып табылатын дендрит клеткалары арқылы интерферон-гамма өнімдерін сақтау қажет.
N-ацетилцистеин немесе NAC глутатионның прекурсоры деп аталатын, сондай-ақ еркін радикалды тазартқыш антиоксидант ретінде пайдаланылатын өте қуатты жасушалық антиоксидант болып табылады. Оның рөлі үшін әдетте танылған ацетаминофеннің уыттылығын жоғалту, NAC, немесе N-ацетил-цистеиннің денсаулық пен сауықтырудың бірнеше артықшылықтары бар екендігі дәлелденді. Сәйкес Cell Journal, NAC сау қабынуға қарсы әрекет етуді қолдауға көмектеседі және адами мерзімге және ерте еңбекке оң әсерін тигізуі мүмкін. Зерттеудің нәтижелері бойынша, ерте босанған және бактериальды вагинозы бар әйелдерде тәулігіне ХННМХ грамм NAC күніне 0.6 аптасынан кейін прогестеронмен бірге ауызша түрде ауызша қабылданады, бұл алдын-ала туудың қайталануына және жаңа туған нәрестелерді жақсартуға қарсы. Қорытындылай келе, NAC-дің бұлшықет құрылысына оң әсері анықталды. Үш минуттан кейін үзіліссіз үзілістен кейін, NAC бұлшықет құрылысын жақсартуда және жұмыс кезінде жалпы шаршауды азайтуда қаншалықты маңызды рөл атқаратынын көрсетеді.
Зерттеушілер сонымен қатар NAC немесе N-ацетил-цистеиннің аналық безінің поликистоз синдромы немесе PCOS бар адамдарға пайда әкелетіндігін анықтады. PCOS, немесе поликистозды аналық без синдромы, репродуктивті жастағы әйелдердің шамамен 5-10 пайызына әсер ететін эндокриндік бездерге қатысты кең таралған ауру. Мұндай пациенттерде метаболикалық синдромның пайда болу қаупі жоғары, мұнда НАК қолдану инсулиннің сау деңгейі мен сезімталдығын қалпына келтіруге көмектесті.
Доктор Алекс Хименездің түсінігі
Глутатион жалпы денсаулық пен денсаулыққа қол жеткізуде және оны сақтауда оның негізгі рөлі арқасында «антиоксиданттар шебері» деп аталды. Адам ағзасы өзінің глютатионын, нашар тамақтануын, ластануын, токсиндерін, есірткі мен / немесе дәрі-дәрмектерді артық пайдалануын, стрессті, жарақаттануды, қартаюды, ауруды және радиацияны шығаруға қабілетті болғанымен, біздің глутатионның табиғи деңгейін төмендете алады. Бұл, өз кезегінде, адамдарға тотығу стресінен, еркін радикалдардан, инфекциялардан және қатерлі ісікпен клеткалық зақымдануға сезімтал болады. Сондықтан глутатионның қосылуы адам ағзасына үлкен пайда әкелуі мүмкін. Широпрактикалық күтім сияқты альтернативті емдеу әдістерімен бірге, глутатион деңгейлері жақсылықты жақсарту үшін тағы да реттелуі мүмкін.
Сонымен қатар, медицина қызметкерлері глутатионды қосымша енгізуді басқа балама емдеу әдістерімен бірге қолдануды ұсынды, мысалы: широпротикалық күтім, жалпы денсаулық пен денсаулықты жақсарту үшін. Антиоксиданттар максималды әл-ауқаттың сақталуына, сондай-ақ жасуша зиянына немесе зақымына әкелетін бос радикалдардың тізбекті реакциясын тежеуге маңызды. Жоғарыда айтылғанындай, глутатион тәрізді күшті антиоксиданттар, сайып келгенде, осы еркін радикалдардың дамуын реттеуге және иммундық жүйенің тиімді жауап беруіне көмектеседі. Зерттеулердің нәтижесі осы широпротикалық күтім сонымен бірге адам ағзасындағы антиоксиданттар белсенділігін арттыра отырып, осы процесте маңызды рөл атқара алады. Широпрактикалық күтім - бұл адам ағзасына дәрі-дәрмектерді және / немесе хирургиялық араласуды қолданбай табиғи емдеуге мүмкіндік беру үшін омыртқаның түзетулерін және манипуляцияларды қолдана отырып, жұлын теңестірулерін түзету үшін немесе емдеудің тиімді әдісі.
Сонымен, антиоксиданттар өздерінің биологиялық қасиеттерін денсаулыққа пайдасы зор екендігін көрсетеді, бұл қазіргі заманғы әлемдегі жасушалардың зақымдануына және / немесе зақымдануына ықпал ететін стресстің, аурулардың және ластанулардың күшейе түсуіне байланысты болуы мүмкін. . Глутатион және оның прекурсоры, NAC немесе N-ацетил-цистеин, антиоксиданттар аймағында өзінің күшті мәртебесін көрсетеді. Хиропрактикалық күтім сияқты баламалы емдеу нұсқаларымен бірге адамдар осы күшті антиоксидант ұсынатын барлық артықшылықтарды пайдалана алады. Біздің ақпарат шеңберіміз хиропрактикамен, сондай-ақ жұлын жарақаттары мен жағдайларымен шектеледі. Тақырыпты талқылау үшін, доктор Хименеске хабарласыңыз немесе бізбен байланысыңыз 915-850-0900 .
Доктор Алекс Хименес мырзаның жетекшілігімен
Қосымша тақырыптар: Back Pain
Арқа ауруы мүгедектіктің ең көп таралған себептерінің бірі және бүкіл әлем бойынша жұмыстан босатылған күндер. Іс жүзінде ауырсыну дәрігерге барудың екінші ең көп тараған себебі ретінде анықталды, бұл тек жоғарғы тыныс жолдарының жұқпалы ауруларымен ғана шектеледі. Шамамен 80 пайызы өмір сүру кезеңінде кем дегенде бір рет арқа ауруы кез-келген түрін сезінеді. Омыртқасы - бұл жұмсақ маталардан тұратын сүйек, буын, байлам және бұлшықеттерден тұратын күрделі құрылым. Осыған байланысты жарақаттар және / немесе ауырлататын жағдайлар сияқты қатты дискілер, ақыр соңында арқа ауырсыну белгілеріне алып келуі мүмкін. Спорттық жарақаттар немесе автомобиль апатының жарақаттары көбінесе арқадағы ауырсынудың жиі себебі болып табылады, бірақ кейде ең қарапайым қозғалыстың ауыр нәтижесі болуы мүмкін. Бақытымызға орай, широпрактикалық күтім сияқты альтернативті емдеу параметрлері омыртқаның түзетуін және манипуляцияларды қолдану арқылы ауырсынуды жеңілдетуге көмектеседі, ақыр соңында ауруды жеңілдетеді.
Ғылыми негізделген хиропрактор доктор Александр Хименес қарайды тотықтырғыш стресс, бұл не, бұл жағдайды түзету үшін денеге және антиоксиданттық қорғанысқа қалай әсер етеді.
Аннотация: Оттегінің реактивті түрлері (ROS) қалыпты жасушалық метаболизм және ауаны ластаушы заттар немесе темекі түтіні сияқты қоршаған орта факторларының нәтижесінде тірі организмдермен түзіледі. ROS жоғары реактивті молекулалар болып табылады және көмірсулар, нуклеин қышқылдары, липидтер және ақуыздар сияқты жасуша құрылымдарын зақымдауы және олардың функцияларын өзгертуі мүмкін. Тотықтырғыштар мен антиоксиданттар арасындағы тепе-теңдіктің тотықтырғыштардың пайдасына ауысуы «тотықтырғыш стресс» деп аталады. Тотықсыздандырғыш және тотықтырғыш (тотықсыздандырғыш) күйдің реттелуі жасушаның өміршеңдігі, активтенуі, пролиферациясы және органның қызметі үшін маңызды. Аэробты организмдердің біріктірілген антиоксиданттық жүйелері бар, олар ферментативті және ферментативті емес антиоксиданттарды қамтиды, олар әдетте ROS зиянды әсерлерін блоктауда тиімді. Дегенмен, патологиялық жағдайларда антиоксиданттық жүйелер шамадан тыс болуы мүмкін. Тотығу стрессі қатерлі ісік, неврологиялық бұзылулар, атеросклероз, гипертония, ишемия/перфузия, қант диабеті, жедел респираторлық дистресс синдромы, идиопатиялық өкпе фиброзы, өкпенің созылмалы обструктивті ауруы және демікпе сияқты көптеген патологиялық жағдайлар мен ауруларға ықпал етеді. Бұл шолуда біз жасушалық тотықтырғыш және антиоксиданттық жүйелерді қорытындылаймыз және тотығу стрессінің жасушалық әсерлері мен механизмдерін талқылаймыз.
Негізгі сөздер: антиоксидант, тотықтырғыш, тотығу стрессі, оттегінің реактивті түрлері, тотығу-тотықсыздану
(WAO журналы 2012; 5:9-19)
Оттегінің реактивті түрлері (ROS) қалыпты жасушалық метаболизм нәтижесінде тірі организмдермен түзіледі. Төмен және орташа концентрацияларда олар жасушаның физиологиялық процестерінде қызмет етеді, бірақ жоғары концентрацияларда олар липидтер, белоктар және ДНҚ сияқты жасуша компоненттеріне жағымсыз өзгерістер тудырады.1�6 Тотықтырғыш/антиоксидант арасындағы тепе-теңдіктің тотықтырғыштардың пайдасына ауысуы Тотығу стрессі қатерлі ісік, неврологиялық бұзылулар, 7-10 атеросклероз, гипертония, ишемия/перфузия, 11 қант диабеті, жедел респираторлық дистресс синдромы, өкпенің идиопатиялық фиброзы, өкпенің созылмалы обструктивті ауруы сияқты көптеген патологиялық жағдайларға ықпал етеді. ,14 және демікпе.15�16 Аэробты организмдердің біріктірілген антиоксиданттық жүйелері бар, олар әдетте ROS зиянды әсерлерін блоктауға тиімді ферменттік және ферментативті емес антиоксиданттарды қамтиды. Дегенмен, патологиялық жағдайларда антиоксиданттық жүйелер шамадан тыс болуы мүмкін. Бұл шолуда біз жасушалық тотықтырғыш және антиоксиданттық жүйелерді және денсаулық және ауру күйіндегі қалпына келтіру және тотықтырғыш (тотықсыздану) күйін реттеуді қорытындылаймыз.
ТОТЫҚТАНДЫРУЛАР
ROS эндогендік көздері
ROS қалыпты жасушалық метаболизм нәтижесінде молекулалық оттегіден түзіледі. ROS екі топқа бөлуге болады: бос радикалдар және радикалды емес. Құрамында бір немесе бірнеше жұпталмаған электрондары бар және сол арқылы молекулаға реактивтілік беретін молекулалар бос радикалдар деп аталады. 2 бос радикал жұпталмаған электрондарын бөліскенде, радикалды емес формалар пайда болады. Физиологиялық маңызы бар 2 негізгі ROS супероксид анионы (O3.), гидроксил радикалы (OH) және сутегі асқын тотығы (H22O2). ROS 2-кестеде жинақталған.
Супероксид анионы молекулалық оттегіге 1 электронның қосылуы арқылы түзіледі.22 Бұл процесс никотин-адениндинуклеотид фосфаты [NAD(P)H] оксидазасы немесе ксантиноксидаза немесе митохондриялық электронды тасымалдау жүйесі арқылы жүзеге асады. Супероксид анионын өндірудің негізгі орны - митохондрия, аденозинтрифосфатты өндіруге арналған жасуша механизмі. Әдетте электрондар оттегінің суға дейін тотықсыздануы үшін митохондриялық электронды тасымалдау тізбегі арқылы тасымалданады, бірақ барлық электрондардың шамамен 1-3% жүйеден ағып, супероксид түзеді. NAD(P)H оксидаза полиморфты ядролық лейкоциттерде, моноциттерде және макрофагтарда кездеседі. Фагоцитоз кезінде бұл жасушалар бактерицидтік белсенділікке әкелетін супероксидтің жарылуын тудырады. Супероксид супероксидті дисмутазалардың әсерінен сутегі асқын тотығына айналады (SODs, EC 1.15.1.1). Сутегі асқын тотығы плазмалық мембрана арқылы оңай таралады. Сутегі асқын тотығы сонымен қатар ксантиноксидаза, аминқышқыл оксидаза және NAD(P)H оксидаза23,24 және пероксисомаларда метаболикалық реакцияларда молекулалық оттегін тұтыну арқылы өндіріледі. ХабереВейс және Фентон реакциялары деп аталатын реакциялар қатарынан H2O2 Fe2 немесе Cu21 сияқты өткізгіш металдардың қатысуымен OH21.25-ге дейін ыдырайды.
Fe31 +�.O2�?Fe2 +�O2 Haber Weiss
Fe2 +�H2O2�?Fe3 +�OH�+ .OH Фентон реакциясы
O 2 өзі де H2 O2-мен әрекеттесіп, OH түзе алады.26,27 Гидроксильді радикал ROS ішіндегі ең реактивті болып табылады және белоктарды, липидтерді, көмірсуларды және ДНҚ-ны зақымдауы мүмкін. Ол сондай-ақ полиқанықпаған май қышқылдарынан электрон алу арқылы липидтердің асқын тотығуын бастай алады.
Гранулоцитарлы ферменттер эозинофил пероксидаза және миелопероксидаза (МПО) арқылы H2O2 реактивтілігін одан әрі кеңейтеді. Белсендірілген нейтрофилдерде H2O2 MPO арқылы тұтынылады. Хлорид ионының қатысуымен H2O2 гипохлор қышқылына (HOCl) айналады. HOCl жоғары тотығу қабілетіне ие және тыныс алу жолдарындағы қоздырғыштарды өлтіруде маңызды рөл атқарады.28 Дегенмен, HOCl сонымен қатар ДНҚ-мен әрекеттесіп, ДНҚ-ақуыздың өзара әрекеттесуін индукциялай алады және пиримидиндік тотығу өнімдерін өндіріп, ДНҚ негіздеріне хлорид қоса алады.29,30 Эозинофил пероксидазасы. және MPO сонымен қатар белоктарды галогендеу, нитрлеу және тирозил радикалдары арқылы протеиннің айқаспалы байланыстары арқылы модификациялау арқылы тотығу стрессіне ықпал етеді.31�33
Басқа оттегіден алынған бос радикалдар - пероксил радикалдары (ROO$). Бұл радикалдардың қарапайым түрі гидропероксил радикалы (HOO$ ) және май қышқылының асқын тотығуында рөл атқарады. Бос радикалдар бүйірлік тізбектегі метилен көміртегінен сутегі атомын алу арқылы липидтердің асқын тотығу тізбегі реакцияларын тудыруы мүмкін. Содан кейін липидті радикал оттегімен әрекеттесіп, пероксил радикалын түзеді. Пероксил радикалы тізбекті реакцияны бастайды және полиқанықпаған май қышқылдарын липидті гидропероксидтерге айналдырады. Липидтердің гидропероксидтері өте тұрақсыз және альдегидтер (мысалы, 4-гидрокси-2,3-ноненал) және малондиальдегидтер (МДҚ) сияқты қайталама өнімдерге оңай ыдырайды. Изопростандар арахидон қышқылының асқын тотығуы арқылы түзілетін липидтердің асқын тотығу өнімдерінің тағы бір тобы болып табылады, сонымен қатар астматикалық науқастардың плазмалық және тыныс конденсаттарында жоғарылайтыны анықталды.34,35 Липидтердің асқын тотығуы жасуша мембраналарының тұтастығын бұзады және олардың қайта орналасуына әкеледі. мембрана құрылымы.
Стандартталған талдаулар арқылы тотығу биомаркерлері ретінде сутегі асқын тотығы, супероксид радикалы, тотыққан глутатион (GSSG), MDAs, изопростандар, карбонилдер және нитротирозинді плазмадан, қаннан немесе бронхоальвеолярлы шаю үлгілерінен оңай өлшеуге болады.
Тотықтырғыштардың экзогендік көзі
Темекі түтіні
Темекі түтінінде көптеген тотықтырғыштар мен бос радикалдар және супероксид және азот оксиді сияқты органикалық қосылыстар бар.36 Сонымен қатар, темекі түтінінің өкпеге ингаляциясы нейтрофилдер мен макрофагтардың жиналуы сияқты кейбір эндогендік механизмдерді белсендіреді, бұл тотықтырғыш зақымдануды одан әрі арттырады. .
Озон экспозициясы
Озонның әсері липидтердің асқын тотығуын тудыруы және нейтрофилдердің тыныс жолдарының эпителийіне түсуін тудыруы мүмкін. Озонға қысқа мерзімді әсер ету сонымен қатар MPO, эозинофильдік катиондық ақуыздар, сондай-ақ лактатдегидрогеназа және альбумин сияқты қабыну медиаторларының бөлінуіне себеп болады.37 Тіпті дені сау адамдарда озон әсері өкпе функцияларының төмендеуіне әкеледі.38 Cho et al39 бөлшектер (ауада ілінген қатты бөлшектер мен сұйық тамшылардың қоспасы) оттегінің тотықсыздануын катализдейді.
Гипероксия
Гипероксия өкпедегі немесе басқа дене тіндеріндегі оттегінің қалыпты парциалды қысымынан жоғары оттегі деңгейінің жағдайын білдіреді. Ол реактивті оттегі мен азот түрлерінің көбірек өндірілуіне әкеледі.40,41
Иондаушы радиация
Иондаушы сәулелену, O2 қатысында гидроксил радикалын, супероксидті және органикалық радикалдарды сутегі асқын тотығы мен органикалық гидропероксидтерге айналдырады. Бұл гидропероксид түрлері Фентон реакциялары арқылы Fe және Cu сияқты тотықсыздандырғыш белсенді металл иондарымен әрекеттеседі және осылайша тотығу стрессін тудырады.42,43 Narayanan және т.б.44 альфа бөлшектерінің әсеріне ұшыраған фибробласттарда жасушаішілік O2 2. және H2O2 айтарлықтай жоғарылағанын көрсетті. плазмалық мембранамен байланысқан NADPH оксидаза арқылы өндіріледі.44 Жасушадан тыс сигналмен реттелетін киназа 1 және 2 (ERK1/2), c-Jun N-терминалды киназа (JNK) және p38 сияқты сигналды тасымалдау молекулалары және транскрипция факторлары, мысалы, белсендіруші ақуыз-1 (AP-1), ядролық фактор-кБ (NF-kB) және p53 белсендіріледі, бұл радиациялық реакцияға байланысты гендер экспрессиясына әкеледі.45�50 Ультракүлгін А (UVA) фотондары тотығу реакцияларын тудырады. порфириндер, NADPH оксидаза және рибофлавиндер сияқты эндогендік фотосенсибилизаторларды қозу арқылы. 8-Оксо-7,8-дигидрогуанин (8-оксоГуа) ОН радикалының, 1-электронды тотықтырғыштардың және негізінен гуанинмен әрекеттесетін синглет оттегінің тотығуы нәтижесінде түзілетін негізгі УКА-делдалдық ДНҚ тотығу өнімі.51 Гуаниннің түзілуі. Оқшауланған ДНҚ-дағы радикал катион иондаушы сәулеленудің тікелей әсері арқылы тиімді пайда болатыны көрсетілді.52,53 Иондаушы сәулеленудің әсерінен кейін глутатионның жасушаішілік деңгейі (GSH) қысқа мерзімге төмендейді, бірақ кейін қайтадан артады.54
Ауыр металл иондары
Темір, мыс, кадмий, сынап, никель, қорғасын және мышьяк сияқты ауыр металл иондары липидтердің асқын тотығуы және ядролық ақуыздармен және ДНҚ-мен реакциясы арқылы ферменттердің белсенділігінің төмендеуі арқылы реактивті радикалдардың түзілуін тудыруы мүмкін.55
Металл арқылы бос радикалдардың пайда болуының маңызды механизмдерінің бірі фентон типті реакция арқылы жүзеге асады. Супероксид ионы мен сутегі асқын тотығы темір және мыс сияқты өтпелі металдармен OH радикалдарын түзу үшін металл катализделген ХаберуВейс/Фентон реакциясы арқылы әрекеттесе алады.
Фентон және Хабер-Вейс типті механизмдерден басқа, белгілі бір металл иондары тиол радикалдары сияқты бос радикалдарды тудыру немесе жасуша сигнал беру жолдарын индукциялау үшін жасушалық молекулалармен тікелей әрекеттесе алады. Бұл радикалдар басқа тиол молекулаларымен де әрекеттесіп, O22.. O22 түзе алады. H2O2-ге айналады, бұл қосымша оттегі радикалының түзілуін тудырады. Кейбір металдар, мысалы, арсенит, жасушалардағы радикалды өндіруші жүйелерді белсендіру арқылы жанама түрде ROS түзілуін индукциялайды.56
Мышьяк – супероксид (O2 2), синглет оттегі (1O2), пероксидтік радикал (ROO), азот оксиді (NO), сутегі асқын тотығы (H2O2) және диметиларсинді пероксил радикалдарын қоса алғанда, әртүрлі ROS түзетін өте улы элемент. CH3)2AsOO ].57�59 Мышьяк (III) қосылыстары антиоксиданттық ферменттерді, әсіресе глутатион-S-трансферазалар (GSTs), глутатион пероксидаза (GSH-Px) және GSH редуктазасы сияқты GSH-тәуелді ферменттерді байланыстыру арқылы тежей алады. - олардың сульфгидрилдік (�SH) топтарына ілесу.60,61
Қорғасын липидтердің асқын тотығуын арттырады.62 Қорғасын әсерінен кейін тіндердің SOD және ми GPx белсенділігінің айтарлықтай төмендеуі хабарланды.63,64 Көптеген ферменттер үшін кофактор ретінде қызмет ететін мырыштың қорғасынмен алмастырылуы мұндай ферменттердің инактивациясына әкеледі. Қорғасынның әсері тіндік тиолдарға әсер ету арқылы GST тежелуіне әкелуі мүмкін.
Металл-катализделген реакциялар нәтижесінде түзілетін ROS ДНҚ негіздерін өзгерте алады. Fe21, Cu21 және Ni21 сияқты металл иондарының тотығу зақымдануы нәтижесінде үш негізді алмастыру, G / C, G / T және C / T болуы мүмкін. Reid et al65 G/C негізінен Fe21 өндіретінін, ал C/T алмастыру Cu21 және Ni21 екенін көрсетті.
АНТИОКСИДАНТТАР
Адам ағзасы тотықтырғыштардың әсерін теңестіруге қызмет ететін әртүрлі антиоксиданттармен жабдықталған. Барлық практикалық мақсаттар үшін оларды 2 санатқа бөлуге болады: ферментативті (2-кесте) және ферментативті емес (3-кесте).
Ферменттік антиоксиданттар
Өкпенің негізгі ферментативті антиоксиданттары - SODs (EC 1.15.1.11), каталаза (EC 1.11.1.6) және GSH-Px (EC 1.11.1.9). Осы негізгі ферменттерге қосымша, басқа антиоксиданттар, соның ішінде гемоксигеназа-1 (EC 1.14.99.3) және тиоредоксиндер (TRXs, EC 1.8.4.10), пероксиредоксиндер (PRXs, EC 1.11.1.15) және глютаредоксиндер сияқты тотықсыздандырғыш ақуыздар , сонымен қатар өкпенің антиоксиданттық қорғанысында маңызды рөл атқаратыны анықталды.
Супероксид әртүрлі көздерден алынатын негізгі ROS болғандықтан, оны SOD арқылы дисмутациялау әрбір жасуша үшін маңызды болып табылады. SOD-тің барлық 3 формасы, яғни CuZn- SOD, Mn-SOD және EC-SOD адамның өкпесінде кеңінен таралған. Mn-SOD митохондрия матрицасында локализацияланған. EC-SOD негізінен жасушадан тыс матрицада, әсіресе I типті коллаген талшықтарының көп мөлшері бар аймақтарда және өкпе және жүйелі тамырлардың айналасында локализацияланған. Ол сондай-ақ бронх эпителийінде, альвеолярлы эпителийде және альвеолярлы макрофагтарда анықталды.66,67 Жалпы алғанда, CuZn-SOD және Mn-SOD әдетте супероксид радикалдарын жаппай жоюшы ретінде әрекет етеді деп есептеледі. Өкпедегі салыстырмалы түрде жоғары EC-SOD деңгейі жасушадан тыс матрица компоненттерімен спецификалық байланысы өкпе матрицасын қорғаудың негізгі құрамдас бөлігі болуы мүмкін.68
SOD әсерінен немесе ксантиноксидаза сияқты оксидазалардың әсерінен түзілетін H2O2 каталаза және GSH-Px арқылы суға дейін тотықсызданады. Каталаза 4 бірдей мономерден тұратын тетрамер түрінде болады, олардың әрқайсысының белсенді орнында гем тобы бар. H2O2 ыдырауы каталаза-феррикаталазаның (суға координацияланған темір) және I қосылысының (оттегі атомымен комплексті темір) 2 конформациясы арасындағы конверсия арқылы жүзеге асады. Каталаза сонымен қатар NADPH-ды қалпына келтіретін эквивалент ретінде байланыстырады, себебі ол суға дейін тотықсызданған кезде H2O2 арқылы ферменттің тотығу инактивациясының (II қосылысының түзілуі) алдын алады.69
H2O2 және липидті гидропероксидтердің (мембраналық липидтердің асқын тотығуы нәтижесінде түзілетін) тотықсыздануына жауапты тотығу-тотықсыздану цикліндегі ферменттерге GSH-Pxs кіреді.70 GSH-Pxs - құрамында бірегей амин қышқылы селеноцистеин бар тетрамерлі ферменттер тобы. белсенді орындарды ашады және H2O2 және липидті пероксидтерді сәйкес спирттерге дейін азайту үшін GSH сияқты төмен молекулалық салмақты тиолдарды пайдаланыңыз. Түрлі гендермен кодталған төрт GSH- Px сипатталған: GSH- Px-1 (жасушалық GSH-Px) барлық жерде кездеседі және H2O2 мен май қышқылының асқын тотығын төмендетеді, бірақ эфирленген пероксил липидтерін емес.71 Этерификацияланған липидтер мембранамен байланысқан GSH арқылы тотықсызданады. -Px-4 (фосфолипидті гидропероксид GSH-Px), ол қалпына келтіретін эквиваленттер ретінде бірнеше түрлі төмен молекулалық тиолдарды пайдалана алады. GSH-Px-2 (асқазан-ішек GSH-Px) асқазан-ішек эпителий жасушаларында локализацияланған, онда ол диеталық пероксидтерді азайтуға қызмет етеді.72 GSH-Px-3 (жасушадан тыс GSH-Px) GSH-Px отбасының жалғыз мүшесі болып табылады. жасушадан тыс бөлім және сүтқоректілердегі ең маңызды жасушадан тыс антиоксиданттық ферменттердің бірі болып саналады. Олардың ішінде жасушадан тыс GSH-Px адамның өкпесінде кеңінен зерттеледі.73
Сонымен қатар, H2O2 утилизациясы құрамында тиол бар бірнеше ферменттермен, атап айтқанда, TRXs (TRX1 және TRX2), тиоредоксин редуктазалары (EC 1.8.1.9) (TRRs), PRXs (олар тиоредоксин пероксидазалары) және глутарокстармен тығыз байланысты.
Цитозолда да, митохондрияда да бар екі TRX және TRR адам жасушаларында сипатталған. Өкпеде TRX және TRR бронх және альвеолярлы эпителийде және макрофагтарда көрінеді. Адам жасушаларында алты түрлі PRX табылды, олардың ультрақұрылымдық бөлінуімен ерекшеленеді. Эксперименттік зерттеулер альвеолярлы эпителийді қорғауда PRX VI маңыздылығын анықтады. Адамның өкпесі бронх эпителийінде, альвеолярлы эпителийде және макрофагтарда барлық PRX-терді көрсетеді.75 Жақында PRX V пероксинитрит редуктаза ретінде қызмет ететіні анықталды,76 бұл оның өкпенің ROS-делдалдық зақымдануының дамуында әлеуетті қорғаныс қосылысы ретінде қызмет етуі мүмкін дегенді білдіреді. .77
Бұл антиоксиданттарға ортақ нәрсе - қалпына келтіретін эквивалент ретінде NADPH қажеттілігі. NADPH каталазаны белсенді түрде сақтайды және GSSG-ны GSH-ге, GSH-Pxs үшін қосалқы субстратқа түрлендіретін TRX және GSH редуктаза (EC 1.6.4.2) арқылы кофактор ретінде пайдаланылады. Жасуша ішілік NADPH, өз кезегінде, глюкоза-1-фосфаттың 6-фосфоглюконолактонға айналуы кезінде пентоза-фосфат жолының бірінші және жылдамдығын шектейтін фермент болып табылатын глюкоза-6-фосфатдегидрогеназа арқылы NADP6 редукциясы арқылы түзіледі. NADPH генерациялау арқылы глюкоза-6-фосфатдегидрогеназа цитозолдық GSH буферлеу қабілетінің (GSH/GSSG) маңызды детерминанты болып табылады, сондықтан оны маңызды, реттегіш антиоксиданттық фермент деп санауға болады.78,79
GSTs (EC 2.5.1.18), басқа антиоксиданттық ферменттер тобы қанықпаған альдегидтер, эпоксидтер және гидропероксидтер сияқты қайталама метаболиттерді белсендірмейді. GST-тердің үш негізгі отбасы сипатталған: цитозолдық GST, митохондриялық GST,80,81 және эйкозаноид пен GSH метаболизмінде рөлі бар мембранамен байланысты микросомалық GST.82 Сүтқоректілерде Альфа, Му деп белгіленген цитозолдық GST жеті класы анықталған. Pi, Sigma, Theta, Omega және Zeta.83�86 Стресссіз жағдайларда Mu және Pi класы GST сәйкесінше Ask1 және JNK киназаларымен әрекеттеседі және бұл киназаларды тежейді.87�89 GSTP1-ден диссоциацияланатыны көрсетілген. JNK тотығу стрессіне жауап ретінде.89 GSTP1 сонымен қатар PRX VI-мен физикалық әрекеттеседі және тотыққан ақуыздың глутатионилденуі арқылы PRX ферментінің белсенділігін қалпына келтіруге әкеледі.90
Ферментативті емес антиоксиданттар
Ферментативті емес антиоксиданттарға витаминдер (С және Е дәрумендері), b-каротин, несеп қышқылы және тиолды құрайтын трипептид (Lg-глутамил-L-цистеинил-Л-глицин) сияқты төмен молекулалы қосылыстар жатады. сульфгидрил) тобы.
С дәрумені (аскорбин қышқылы)
Суда еритін С дәрумені (аскорбин қышқылы) оттегінің бос радикалдарын тазарту арқылы жасушаішілік және жасушадан тыс су фазасының антиоксиданттық қабілетін қамтамасыз етеді. Ол Е дәруменінің бос радикалдарын қайтадан Е витаминіне айналдырады. Оның плазмадағы деңгейі жас ұлғайған сайын төмендейтіні көрсетілген.91,92
Е дәрумені (а-токоферол)
Липидтерде еритін Е витамині жасуша мембранасының гидрофобты ішкі аймағында шоғырланған және тотықтырғыш әсер ететін мембрана зақымдануынан негізгі қорғаныс болып табылады. Е витамині липидтердің тотығуы кезінде түзілетін пероксил радикалына электрон береді. а-токоферол Е витаминінің ең белсенді түрі және жасушадағы негізгі мембранамен байланысқан антиоксидант болып табылады. Е витамині қатерлі ісік жасушаларының апоптозын тудырады және бос радикалдардың түзілуін тежейді.93
Глутатион
GSH барлық жасуша бөлімдерінде өте көп және негізгі еритін антиоксидант болып табылады. GSH/GSSG қатынасы тотығу стрессінің негізгі детерминанты болып табылады. GSH өзінің антиоксиданттық әсерін бірнеше жолмен көрсетеді.94 Ол GSH-Px әрекеті арқылы сутегі асқын тотығы мен липидтердің асқын тотығын детоксикациялайды. GSH өзінің электронын H2O2-ге H2O және O2-ге дейін тотықсыздандыру үшін береді. GSSG электрон доноры ретінде NAD(P)H пайдаланатын GSH редуктаза арқылы қайтадан GSH-ға төмендейді. GSH-Pxs жасуша мембранасын липидтердің асқын тотығуынан қорғау үшін де маңызды. Азайған глутатион протондарды мембраналық липидтерге береді және оларды тотықтырғыш әсерлерден қорғайды.95
GSH – GSH-Px және трансфераза сияқты бірнеше детоксикациялаушы ферменттердің кофакторы. Ол С және Е дәрумендерін белсенді түрге айналдыруда маңызды рөл атқарады. GSH проапоптотикалық және антиапоптотикалық сигнал беру жолдарымен әрекеттесу арқылы жасушаларды апоптоздан қорғайды.94 Сондай-ақ ол AP-1, NF-kB және Sp-1 сияқты бірнеше транскрипция факторларын реттейді және белсендіреді.
Каротиноидтар (b-каротин)
Каротиноидтар - өсімдіктерде кездесетін пигменттер. Ең алдымен, b-каротиннің пероксил (ROO), гидроксил (OH) және супероксид (O22.) радикалдарымен әрекеттесетіні анықталды.96 Каротиноидтар өздерінің антиоксиданттық әсерін төмен оттегінің парциалды қысымында көрсетеді, бірақ жоғары оттегіде прототықтырғыш әсерлері болуы мүмкін. концентрациялары.97 Каротиноидтар да, ретиноин қышқылдары да (РА) транскрипция факторларын реттеуге қабілетті.98 b-Каротин тотықтырғышпен индукцияланған NF-kB белсенділенуін және интерлейкин (IL)-6 және ісік некрозының факторы-а өндірісін тежейді. Каротиноидтар жасушалардың апоптозына да әсер етеді. РА антипролиферативті әсерлері бірнеше зерттеулерде көрсетілген. РА-ның бұл әсері негізінен ретиной қышқылының рецепторлары арқылы жүзеге асады және жасуша түрлеріне байланысты өзгереді. Сүт безінің карциномасының жасушаларында ретиной қышқылы рецепторының жасушалық циклді тоқтату, апоптоз немесе екеуін де индукциялау арқылы өсудің тежелуін қоздыратыны көрсетілді.99,100
ТОТЫҚТЫРУ СТРЕССІНІҢ ӘСЕРІ: ГЕНЕТИКАЛЫҚ, ФИЗИОЛОГИЯЛЫҚ, ЖӘНЕ БИОХИМИЯЛЫҚ МЕХАНИЗМДЕР
Тотығу стрессі антиоксиданттар мен ROS арасындағы тепе-теңдік антиоксиданттардың азаюына немесе ROS жинақталуына байланысты бұзылған кезде пайда болады. Тотығу стрессі орын алған кезде жасушалар қорғаныс ферменттерін, транскрипция факторларын және құрылымдық ақуыздарды кодтайтын гендерді белсендіру немесе өшіру арқылы тотығу әсерлеріне қарсы тұруға және тотығу-тотықсыздану тепе-теңдігін қалпына келтіруге тырысады.101,102 Тотыққан және тотықсызданған глутатион (2GSH/GSSG) арасындағы қатынас бір. организмдегі тотығу стресінің маңызды детерминанттары. Денедегі ROS жоғары өндірісі ДНҚ құрылымын өзгертуі мүмкін, нәтижесінде белоктар мен липидтердің модификациясы, бірнеше стресстен туындаған транскрипция факторларының белсендірілуі және қабынуға қарсы және қабынуға қарсы цитокиндердің өндірілуі мүмкін.
Тотығу стресінің ДНҚ-ға әсері
ROS бірнеше жолмен ДНҚ модификацияларына әкелуі мүмкін, ол негіздердің ыдырауын, бір немесе екі тізбекті ДНҚ үзілістерін, пуринді, пиримидинді немесе қантпен байланысты модификацияларды, мутацияларды, делецияларды немесе транслокацияларды және ақуыздармен айқаспалы байланысты қамтиды. Осы ДНҚ модификацияларының көпшілігі (1-сурет) канцерогенезге, қартаюға және нейродегенеративті, жүрек-қан тамырлары және аутоиммунды ауруларға өте маңызды. Темекі түтіні, тотығу-тотықсыздандырылған металдар және темір, кадмий, хром және мышьяк сияқты тотықсыздандырғыш емес металдар да бос радикалдарды тудыру немесе тиол топтарымен байланысу арқылы канцерогенезге және қартаюға қатысады. 8-OH-G түзілуі тотығу стрессі арқылы пайда болатын ең белгілі ДНҚ зақымдануы болып табылады және канцерогенездің ықтимал биомаркері болып табылады.
Гендердің промоутер аймақтарында транскрипция факторлары үшін консенсус тізбегі бар. Бұл транскрипция факторын байланыстыратын тораптар тотықтырғыш шабуылдарға сезімтал GC-қа бай тізбектерді қамтиды. Транскрипция факторының байланысу орындарында 8-OH-G ДНҚ түзілуі транскрипция факторларының байланысуын өзгерте алады және осылайша AP-1 және Sp-1 мақсатты тізбектері үшін көрсетілгендей байланысты гендердің экспрессиясын өзгерте алады.103 8-OH-G-ден басқа, 8,59 -цикло-29 -дезоксиаденозин (цикло-дА) егер TATA қорабында орналасқан болса, жасуша жүйесіндегі репортер генінен транскрипцияны тежейтіні де көрсетілген.104 TATA-байланыстырушы ақуыз ДНҚ иілуін өзгерту арқылы транскрипцияны бастайды. . TATA-байланыстыратын ақуыздың байланысуы цикло-дА қатысуымен бұзылуы мүмкін.
Тотығу стрессі микросателлиттік (қысқа тандемді қайталаулар) аймақтарының тұрақсыздығын тудырады. Тотығу-тотықсыздандырғыш белсенді металл иондары, гидроксил радикалдары микросателлиттің тұрақсыздығын арттырады.105 Тотықтырғыштың зақымдануынан туындаған бір тізбекті ДНҚ үзілістерін жасушалар оңай көтеретін болса да, иондаушы сәулеленуден туындаған қос тізбекті ДНҚ үзілістері жасушаның тіршілігіне айтарлықтай қауіп төндіруі мүмкін.106
ДНҚ-дағы CpG аралдарында метилдену маңызды эпигенетикалық механизм болып табылады, ол геннің үнсіздігіне әкелуі мүмкін. 5-MeCyt-тің 5-гидроксиметил урацилге (5-OHMeUra) дейін тотығуы тимин немесе 5-гидроксиметил цитозин аралық өнімдерінің дезаминдену/тотығу реакциялары арқылы жүзеге асуы мүмкін.107 Модуляциялаушы ген экспрессиясынан басқа, ДНҚ метилденуі хроматин ұйымына да әсер ететін сияқты.108 Тотығу шабуылдарынан туындаған аберрантты ДНҚ метилдену үлгілері ДНҚ жөндеу белсенділігіне де әсер етеді.
Липидтерге тотығу стресінің әсері
ROS липидтердің асқын тотығуын индукциялауы және мембранамен байланысқан рецепторлар мен ферменттерді инактивациялауы және тіндердің өткізгіштігін жоғарылатуы мүмкін мембрананың липидті қос қабатының орналасуын бұзуы мүмкін.109 MDA және қанықпаған альдегидтер сияқты липидтердің асқын тотығу өнімдері ақуыз кроссін түзу арқылы көптеген жасушалық ақуыздарды инактивациялауға қабілетті. -байланыстар.110�112 4-Гидрокси-2-ноненальді жасушаішілік GSH таусылуын тудырады және пероксид өндірісін индукциялайды, 113,114 эпидермиялық өсу факторы рецепторын белсендіреді,115 және фибронектин өндірісін индукциялайды. , тотығу стрессінің жанама биомаркерлері ретінде пайдаланылды және созылмалы обструктивті өкпе ауруы бар науқастардың немесе темекі шегетіндердің дем шығаратын конденсатында немесе бронхоальвеолярлы шаю сұйықтығында немесе өкпеде жоғары деңгейлер көрсетілді.116-117
Ақуыздарға тотығу стресінің әсері
ROS пептидтік тізбектің фрагментациясын, белоктардың электр зарядының өзгеруін, белоктардың айқаспалы байланысын және нақты аминқышқылдарының тотығуын тудыруы мүмкін, сондықтан арнайы протеазалардың ыдырауы арқылы протеолизге сезімталдықтың жоғарылауына әкеледі.120 Ақуыздардағы цистеин мен метионин қалдықтары. тотығуға әсіресе сезімтал.121 Белоктардың сульфгидрильді топтарының немесе метионин қалдықтарының тотығуы конформациялық өзгерістерге, ақуыздың ашылуына және ыдырауына әкеледі.8,121�123 Белсенді жерлерінде немесе оларға жақын металдары бар ферменттер металдың катализделген тотығуына әсіресе сезімтал. Ферменттердің тотығу модификациясы олардың белсенділігін тежейтіні көрсетілген.124,125
Кейбір жағдайларда белоктардың спецификалық тотығуы орын алуы мүмкін. Мысалы, метионинді метионин сульфоксиді126 және фенилаланинді о-тирозинге127 тотықтыруға болады; сульфгидрильді топтар дисульфидтік байланыстар түзу үшін тотығуы мүмкін;128 және карбонил топтары белоктардың бүйірлік тізбегіне енуі мүмкін. Гамма сәулелері, металл катализделген тотығу, HOCl және озон карбонил топтарының түзілуіне себеп болуы мүмкін.129
Сигналдың өтуіне тотығу кернеуінің әсері
ROS сигналды тасымалдауға қатысатын бірнеше гендердің экспрессиясын тудыруы мүмкін.1,130 GSH/GSSG үшін жоғары қатынас жасушаны тотығу зақымдануынан қорғау үшін маңызды. Бұл арақатынастың бұзылуы қабыну реакциясына қатысатын NF-kB, AP-1, белсендірілген Т жасушаларының ядролық факторы және гипоксия-индукцияланатын фактор 1 сияқты тотығу-тотықсыздануға сезімтал транскрипция факторларының белсендірілуін тудырады. ROS арқылы транскрипция факторларының белсендірілуі ақпаратты жасушаның сыртынан ішкі жағына тасымалдайтын сигнал беру каскадтары арқылы жүзеге асырылады. Тирозинкиназа рецепторлары, өсу факторы рецепторларының көпшілігі, мысалы, эпидермиялық өсу факторы рецепторы, тамырлық эндотелий өсу факторы рецепторлары және тромбоциттерден алынған өсу факторы үшін рецепторлар, ақуыз тирозин фосфатазалары және серин/треонинкиназалар ROS нысанасы болып табылады.131-133 Митогенмен белсендірілген протеинкиназалар отбасының мүшелері болып табылатын және пролиферация, дифференциация және апоптозды қоса алғанда жасушадағы бірнеше процестерге қатысатын жасушадан тыс сигналмен реттелетін киназалар, JNK және p38 тотықтырғыштармен реттелуі мүмкін.
Тотығу стресс жағдайында c-Jun ДНҚ-байланыстыру орнындағы цистеин қалдықтары, кейбір AP-1 суббірліктер және ингибиторлық кБ киназа қайтымды S-глутатиоляцияға ұшырайды. Глутаредоксин мен TRX NF-kB және AP-1, p38 митогенмен белсендірілген протеинкиназа және JNK.134�137 сияқты тотығу-тотықсыздануға сезімтал сигналдық жолдарды реттеуде маңызды рөл атқаратыны хабарланды.
NF-kB ROS, бос радикалдар және ультракүлгін сәулелену сияқты тотығу стресс жағдайларына жауап ретінде белсендірілуі мүмкін.138 IkB фосфорлануы NF-kB босатады және оның ген транскрипциясын белсендіру үшін ядроға енуіне мүмкіндік береді.139 Бірқатар киназалар бар серин қалдықтарында IkBs фосфорланатыны хабарланды. Бұл киназалар NF-kB белсендіруіне арналған тотығу сигналдарының нысанасы болып табылады.140 Тотықсыздандырғыштар NF-kB ДНҚ байланысуын күшейтеді, ал тотықтырғыштар NF-kB ДНҚ байланысуын тежейді. TRX NF-kB реттелуінде 2 қарама-қарсы әрекетті орындауы мүмкін: цитоплазмада ол IkB деградациясын блоктайды және NF-kB активациясын тежейді, бірақ ядродағы NF-kB ДНҚ байланысуын күшейтеді.141 Тотығуға байланысты деградация арқылы NF-kB белсендіру IkB антиоксиданттық қорғаныспен байланысты бірнеше гендердің белсендірілуіне әкеледі. NF-kB IL-1b, IL-6, ісік некрозының факторы-a, IL-8 және бірнеше адгезиялық молекулалар сияқты иммундық жауапқа қатысатын бірнеше гендердің экспрессиясын реттейді.142,143 NF-kB сонымен қатар ангиогенезді және пролиферацияны реттейді және жасушалардың дифференциациясы.
AP-1 де тотығу-тотықсыздану күйімен реттеледі. H2O2 болған кезде кейбір металл иондары AP-1 активациясын индукциялауы мүмкін. GSH/GSSG арақатынасының артуы AP-1 байланысуын күшейтеді, ал GSSG AP-1.144 ДНҚ байланысуын тежейді, Fos/Jun гетеродимерінің ДНҚ байланысуы әрқайсысының ДНҚ-байланыстыру доменінде бір ғана сақталған цистеиннің азаюы арқылы артады. ақуыздар145, ал AP-1 ДНҚ байланысуы көптеген жасуша түрлерінде GSSG арқылы тежелуі мүмкін, бұл цистеин қалдықтары арқылы дисульфидтік байланыстың түзілуі AP-1 ДНҚ байланысуын тежейді деп болжайды.146,147 Тотығу стрессі арқылы сигнал беру 2-суретте жинақталған.
ҚОРЫТЫНДЫ
Тотығу стрессі оттегін пайдаланатын және олардың арасындағы тепе-теңдікті өзгертетін метаболикалық реакциялар арқылы ROS шамадан тыс өндірілуінен туындауы мүмкін. тотықтырғыш/антиоксидант тотықтырғыштардың пайдасына жағдай жасайды. ROS жасушалардың метаболизмі және қоршаған орта факторлары, мысалы, ауаны ластаушы заттар немесе темекі түтіні арқылы өндіріледі. ROS жоғары реактивті молекулалар, өйткені олардың құрылымында жұпталмаған электрондар бар және көмірсулар, нуклеин қышқылдары, липидтер және ақуыздар сияқты жасушадағы бірнеше биологиялық макромолекулалармен әрекеттеседі және олардың функцияларын өзгертеді. ROS сонымен қатар тотығу-тотықсыздануға сезімтал транскрипция факторларының жоғарылауы және гистонның ацетилденуі/децетилденуі арқылы хроматиннің қайта құрылуы арқылы бірнеше гендердің экспрессиясына әсер етеді. Тотығу-тотықсыздану күйінің реттелуі жасушаның өміршеңдігі, активтенуі, көбеюі және мүшелер қызметі үшін өте маңызды.
СІЛТЕМЕЛЕР
1. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Тотығу стрессінен туындаған қатерлі ісіктегі бос радикалдар, металдар және антиоксиданттар. Chem Biol әрекеттесуі. 2006;160:1�40.
2. Halliwell B, Gutterridge JMC. Биология мен медицинадағы бос радикалдар. 3-ші басылым. Нью-Йорк: Оксфорд университетінің баспасы; 1999.
3. Марнетт Л.Дж. Липидтердің асқын тотығуы - малондиальдегидпен ДНҚ зақымдануы. Мутат Рес. 1999;424:83�95.
4. Siems WG, Grune T, Esterbauer H. 4-Егеуқұйрықтың жіңішке ішектерінің ишемиялары мен реперфузиясы кезіндегі гидроксиноненальды түзілу. �Өмір ғылымы. 1995;57:785�789.
5. Stadtman ER. Қартаюдағы тотықтырғыш түрлердің рөлі. Curr Med Chem. 2004;11:1105�1112.
6. Ванг М.Й., Дхигра К, Гиттельман В.Н., Лиер Дж.Г., деАндраде М, Ли ДХ. Адамның кеуде тіндеріндегі липидтердің асқын тотығуынан туындаған болжамды малондиальдегидтің ДНҚ қосындылары. Қатерлі ісік эпидемиолын биомаркерлері Алдыңғы. 1996;5:705�710.
7. Дженнер П. Паркинсон ауруындағы тотығу стрессі. Энн Нейроль. 2003;53: S26�S36.
8. Lyras L, Cairns NJ, Jenner A, Jenner P, Halliwell B. Альцгеймер ауруы бар науқастардан мидағы ақуыздардың, липидтердің және ДНҚ-ның тотығу зақымдалуын бағалау. J Neurochem. 1997;68:2061�2069.
9. Сайре Л.М., Смит М.А., Перри Г. Нейродегенеративті аурудағы тотығу стрессінің химиясы және биохимиясы. Curr Med Chem. 2001;8:721�738.
10. Тошнивал П.К., Зарлинг Е.Дж. Көптеген склерозда липидтердің асқын тотығуының жоғарылауының дәлелі. Neurochem Res. 1992;17:205�207.
11. Dhalla NS, Temsah RM, Neticadan T. Жүрек-қан тамырлары ауруларындағы тотығу стрессінің рөлі. J Гипертония. 2000;18:655�673.
12. Каспарова С, Брезова В, Валько М, Хорецки Дж, Млинарик В, т.б. Созылмалы ми гипоперфузиясының егеуқұйрық үлгісіндегі тотығу стрессін зерттеу. Neurochem Int. 2005;46:601�611.
13. Kerr S, Brosnan MJ, McIntyre M, Reid JL, Dominiczak AF, Hamilton CA. Генетикалық гипертензия үлгісінде супероксид анионының өндірісі жоғарылайды: эндотелийдің рөлі. Гипертония. 1999;33:1353�1358.
14. Кукрея RC, Hess ML. Оттегінің бос радикалдық жүйесі: теңдеулерден мембраналық-ақуыздық өзара әрекеттесу арқылы жүрек-қан тамырларының зақымдануы мен қорғанысына дейін. Жүрек-қан тамырлары рес. 1992;26:641�655.
15. Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, т.б. Темекі шегу адам өкпесінің орталық бөлігінде ДНҚ-ның тотығу зақымдалуының, 8-гидроксидеоксигуанозиннің жоғарылауын тудырады. Канцерогенез. 1997;18:1763–1766.
16. Andreadis AA, Hazen SL, Comhair SA, Erzurum SC. Демікпедегі тотығу және нитрозативтік құбылыстар. Free Radic Biol Med. 2003;35:213�225.
17. Comhair SA, Ricci KS, Arroliga M, Lara AR, Dweik RA, т.б. Жүйелі супероксид дисмутаза тапшылығының астмадағы ауа ағынының кедергісіне корреляциясы. Am J Respir Crit Care Med. 2005;172:306�313.
18. Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, т.б. Астматикалық тыныс алу жолдарының ремоделизациясы мен реактивтілігінің патофизиологиясындағы супероксидті дисмутаза инактивациясы. Мен Дж Патхол. 2005;166:663�674.
19. Дут Р, Диздар Е.А., Бирбен Е, Сакесен С, Сойер ОУ, Беслер Т, Калайчи О. Тотығу стрессі және демікпесі бар балалардың тыныс алу жолдарындағы оның детерминанттары. Аллергия. 2008;63:1605�1609.
20. Ercan H, Birben E, Dizdar EA, Keskin O, Karaaslan C, et al. Балалық демікпедегі тотығу стрессі және оксидант жарақатының генетикалық және эпидемиологиялық детерминанттары. J Allergy Clin Immunol. 2006;118:1097�1104.
21. Фицпатрик AM, Teague WG, Holguin F, Yeh M, Brown LA. Ауыр астма зерттеу бағдарламасы. Ауыр демікпесі бар балаларда тыныс алу жолындағы глутатион гомеостазы өзгереді: тотықтырғыш стресстің дәлелі. J Allergy Clin Immunol. 2009;123:146-152.
22. Миллер ДМ, Бьюттнер ГР, Ауст СД. Өтпелі металдар «автототығу» реакцияларының катализаторы ретінде. Free Radic Biol Med. 1990;8:95–108.
23. Dupuy C, Virion A, Ohayon R, Kaniewski J, Dïme D, Pommier J. Қалқанша безінің плазмалық мембранасындағы NADPH оксидазасымен катализделген сутегі асқын тотығының түзілу механизмі. J Biol Chem. 1991;266:3739�3743.
24. Гранджер Д.Н. Ишемия-перфузиялық зақымданудағы ксантиноксидаза мен гранулоциттердің рөлі. Мен J Physiol. 1988;255:H1269�H1275.
25. Fenton HJH. Темірдің қатысуымен шарап қышқылының тотығуы. J Chem Soc. 1984;65:899�910.
26. Хабер Ф, Вайсс Дж. Темір тұздарымен сутегі асқын тотығының каталитикалық ыдырауы. Proc R Soc Lond Ser A. 1934;147:332�351.
27. Лиочев С.И., Фридович И. 70 жылдан кейін Хабер Вайсс циклі: балама көзқарас. Redox Rep. 2002;7:55�57.
28. Клебанов С.Ж. Миелопероксидаза: дос және жау. J Leukoc Biol. 2005;77:598�625.
29. Whiteman M, Jenner A, Halliwell B. Оқшауланған бұзау тимус ДНҚ-дағы гипохлорлы қышқылмен индукцияланған негіз модификациялары. Chem Res Toxicol. 1997;10:1240–1246.
30. Құлшарық ҚБ, Хайнеке Дж.В. Адамның фагоциттерінің миелопероксидаза жүйесі шығаратын гипохлор қышқылы ДНҚ мен ақуыз арасында ковалентті айқаспалы байланыстарды тудырады. Биохимия. 2001;40:3648�3656.
31. Бреннан МЛ, Ву В, Фу Х, Шен З, Сон В және т.б. Екі қарама-қайшылық туралы әңгіме: эозинофил пероксидаза мен миелопероксидаза тапшылығы бар тышқандарды қолдану арқылы in vivo нитротирозин түзудегі пероксидазалардың рөлін және пероксидаза тудыратын реактивті азот түрлерінің табиғатын анықтау. J Biol Chem. 2002;277:17415�17427.
32. Denzler KL, Borchers MT, Crosby JR, Cieslewicz G, Hines EM, т.б. Өкпе қабынуының тінтуірдің сопақ буминдік-шаллендж үлгісінде кең көлемде эозинофилді дегрануляциялау және тыныс алу жолдары ақуыздарының пероксидаза арқылы тотығуы байқалмайды. J Иммунол. 2001;167:1672-1682.
33. Ван Дален Си Дж., Уинтерборн КС, Сентилмохан Р, Кетл А.Дж. Нитрит субстрат және миелопероксидаза ингибиторы ретінде. Қабыну орындарында нитрлеу және гипохлор қышқылының түзілуінің салдары. J Biol Chem. 2000;275:11638�11644.
34. Wood LG, Fitzgerald DA, Gibson PG, Cooper DM, Garg ML. Плазмадағы изопростандармен анықталған липидтердің асқын тотығуы жеңіл демікпедегі аурудың ауырлығына байланысты. Липидтер. 2000;35:967�974.
35. Монтуши П, Корради М, Сиабаттони Г, Найтингейл Дж, Харитонов С.А., Барнс П.Дж. Демікпемен ауыратын науқастардың дем шығару конденсатында тотығу стрессінің белгісі болып табылатын 8-изопростанның жоғарылауы. Am J Respir Crit Care Med. 1999;160:216�220.
36. Шіркеу DF, Pryor WA. Темекі түтінінің еркін радикалды химиясы және оның токсикологиялық салдары. Қоршаған орта денсаулығының перспективасы. 1985;64:111�126.
37. Hiltermann JT, Lapperre TS, van Bree L, Steerenberg PA, Brahim JJ және т.б. Астматиктердің қақырығы мен бронхиалды шаю сұйықтығында бағаланған озоннан туындаған қабыну: ауаның ластануы мен демікпеге арналған эпидемиологиялық зерттеулердегі жаңа инвазивті емес құрал. Free Radic Biol Med. 1999;27:1448–1454.
38. Найтингейл Д.А., Роджерс Д.Ф., Барнс П.Дж. Қалыпты және демікпемен ауыратын адамдарда ингаляциялық озонның деммен шығарылған азот оксидіне, өкпе функциясына және индукцияланған қақырықтыға әсері. Кеуде. 1999;54:1061�1069.
39. Чо А.К., Сиутас С, Мигель А.Х., Кумагай Ю, Шмитц Д.А., т.б. Лос-Анджелес бассейніндегі әртүрлі учаскелердегі ауадағы бөлшектердің тотығу-тотықсыздану белсенділігі. Environ Res. 2005;99:40�47.
40. Comhair SA, Thomassen MJ, Erzurum SC. 2% O(100) немесе темекі түтініне ұшыраған сау адамдардың тыныс алу жолдарында жасушадан тыс глутатион пероксидаза мен азот оксиді синтаза 2 дифференциалды индукциясы. Am J Respir Cell Mol Biol. 2000;23:350�354.
41. Matthay MA, Geiser T, Matalon S, Ischiropoulos H. Жедел респираторлық дистресс синдромындағы оксидантпен әсер ететін өкпе жарақаты. Crit Care Med. 1999;27:2028–2030 ж.
42. Biaglow JE, Mitchell JB, Held K. Пероксид пен супероксидтің рентгендік реакциядағы маңызы. Int J Radiat Oncol Biol Phys. 1992;22:665-669.
43. Чиу С.М., Сюэ Л.И., Фридман Л.Р., Олейник Н.Л. Иондаушы сәулеленуге ядролық матрицаны бекіту орындарын мыс иондары арқылы сенсибилизациялау. Биохимия. 1993;32:6214�6219.
44. Нараянан П.К., Гудвин Е.Х., Лехнерт Б.Е. Альфа бөлшектері адам жасушаларында супероксид аниондары мен сутегі асқын тотығының биологиялық өндірісін бастайды. Cancer Res. 1997;57:3963�3971.
45. Таттл СВ, Варнес М.Е., Митчелл Дж.Б., Биаглоу Дж.Э. Химиялық тотықтырғыштарға және тотығу пентозалық цикл белсенділігінде жетіспейтін CHO жасушаларының радиациясына сезімталдық. Int J Radiat Oncol Biol Phys. 1992;22: 671�675.
46. Гуо Г, Ян-Сандерс Ю, Лин-Кук БД, Ван Т, Тамае Д, т.б. Марганец
радиациялық индукцияда супероксидті дисмутаза арқылы ген экспрессиясы
адаптивті жауаптар. Мол жасушалық биол. 2003;23:2362�2378.
47. Azzam EI, де Толедо SM, Spitz DR, Little JB. Тотығу метаболизмі
сигнал беруді және бақылаушыда микронуклеус түзілуін модуляциялайды
а-бөлшектердің жасушалары адамның қалыпты фибробласттарын сәулелендіріп жіберді. Cancer Res.
2002; 62: 5436 5442.
48. Лич Дж.К., Ван Туйл Г, Лин П.С., Шмидт-Уллрих Р, Миккельсен Р.Б.
Иондаушы сәулелену әсерінен митохондрияға тәуелді реактивті генерация
оттегі/азот. Cancer Res. 2001;61:3894�3901.
49. Dent P, Yacoub A, Fisher PB, Hagan MP, Grant S. MAPK pathways in in
радиациялық реакциялар. Онкоген. 2003;22:5885�5896.
50. Вей СДж, Ботеро А, Хирота К, Брэдбери СМ, Марковина С, т.б. Тиоредоксин
ядролық транслокация және тотықсыздану-1 факторымен әрекеттесу иондаушы сәулеленуге жауап ретінде AP-1 транскрипция факторын белсендіреді. Cancer Res. 2000;60:6688�6695.
51. Кадет Дж, Дуки Т, Гаспарутто Д, Раванат Дж.Л. ДНҚ-ның тотығу зақымдалуы: түзілу, өлшеу және биохимиялық ерекшеліктері. Мутат Рес. 2003;531:5�23.
52. Yokoya A, Cunniffe SM, O�Neill P. Гидратацияның гаммарадиация арқылы плазмидті ДНҚ пленкаларында жіп үзілуі мен негіздік зақымданулардың индукциясына әсері. J Am Chem Soc. 2002;124:8859�8866.
53. Янссен Ю.М., Ван Хоутен Б, Борм П.Дж., Моссман Б.Т. Тотығу зақымдалуына жасуша мен ұлпаның жауаптары. Lab Invest. 1993;69:261�274.
54. Иванага М, Мори К, Иида Т, Урата Ю, Мацуо Т, т.б. Адамның T98G глиобластома жасушаларында иондаушы сәулелену арқылы гамма-глутамилцистеин синтетазасының каппа В ядролық факторына тәуелді индукциясы. Free Radic Biol Med. 1998;24:1256-1268.
55. Stohs SJ, Bagchi D. Металл иондарының уыттылығындағы тотығу механизмдері. Free Radic Biol Med. 1995;18:321�336.
56. Леонард С.С., Харрис Г.К., Ши X. Металдан туындаған тотығу кернеуі және сигналды беру. Free Radic Biol Med. 2004;37:1921–1942.
57. Ши Х, Ши Х, Лю КДж. Мышьяктың уыттылығы мен канцерогенезінің тотығу механизмі. Мол жасушаларының биохимиясы. 2004;255:67�78.
58. Pi J, Horiguchi S, Sun Y, Nikaido M, Shimojo N, Hayashi T. Қояндардағы арсенаттың ұзақ уақыт бойы ауызша әсерінен туындаған азот оксиді түзілуінің бұзылуының ықтимал механизмі. Free Radic Biol Med.2003;35:102�113.
59. Rin K, Kawaguchi K, Yamanaka K, Tezuka M, Oku N, Okada S. Бейорганикалық мышьяк метаболиті диметиларсин қышқылымен индукцияланған ДНҚ тізбегінің үзілуі супероксид анионының радикалдарымен күшті күшейтіледі. Biol Pharm Bull. 1995;18:45�58.
60. Waalkes MP, Liu J, Ward JM, Diwan LA. Мышьяк канцерогенезінің негізінде жатқан механизмдер: жүктілік кезінде бейорганикалық мышьяк әсеріне ұшыраған тышқандардың жоғары сезімталдығы. Токсикология. 2004;198:31�38.
61. Шиллер СМ, Фаулер Б.А., Вудс Дж.С. Мышьяктың пируватдегидрогеназаның активтенуіне әсері. Қоршаған орта денсаулығының перспективасы. 1977;19:205�207.
62. Monterio HP, Bechara EJH, Abdalla DSP. Неврологиялық порфирияларға және қорғасынмен улануға бос радикалдардың қатысуы. Мол жасушаларының биохимиясы. 1991;103:73�83.
63. Tripathi RM, Raghunath R, Mahapatra S. Қандағы қорғасын және оның балалардың Cd, Cu, Zn, Fe және гемоглобин деңгейіне әсері. Sci Total Environ. 2001;277:161�168.
64. Неру Б, Дуа Р. Диеталық селеннің қорғасынның нейроуыттылығына әсері. J Environ Pathol Toxicol Oncol. 1997;16:47-50.
65. Рейд ТМ, Фейг Д.И., Лоеб Л.А. Метал-индукцияланған оттегі радикалдарымен мутагенез. Қоршаған орта денсаулығының перспективасы. 1994;102(3-қосымша):57�61.
66. Кинула ВЛ, Крапо Дж. Өкпе және адамның өкпе аурулары кезіндегі супероксид дисмутазалары. Am J Respir Crit Care Med. 2003;167:1600–1619.
67. Кинула ВЛ. Адамның өкпесіндегі қабыну күйлері кезінде оттегі метаболиттерінің түзілуі және ыдырауы. Curr препараты қабыну аллергиясына бағытталған. 2005;4:465–470.
68. Зелько И.Н., Мариани Т.Дж., Фольц Р.Дж. Супероксидті дисмутаза мультигендік отбасы: CuZn-SOD (SOD1), Mn-SOD (SOD2) және EC-SOD (SOD3) гендік құрылымдарын, эволюциясын және экспрессиясын салыстыру. Free Radic Biol Med. 2002;33:337�349.
69. Киркман Х.Н., Ролфо М, Феррарис А.М., Гаетани Г.Ф. Каталазаны NADPH арқылы қорғау механизмдері. Кинетика және стехиометрия. J Biol Chem. 1999;274:13908�13914.
70. Флохо L. Глутатионпероксидаза. Негізгі өмір ғылымы. 1988;49:663�668.
71. Артур Дж.Р. Глутатион пероксидазалары. Cell Mol Life Sci. 2000;57:1825–1835.
72. Чу ФФ, Доросшоу Дж.Х., Эсворси Р.С. Жаңа жасушалық селенге тәуелді глутатион пероксидазасының GSHPx-GI экспрессиясы, сипаттамасы және тіндік таралуы. J Biol Chem. 1993;268:2571�2576.
73. Comhair SA, Bhathena PR, Farver C, Thunnissen FB, Erzurum SC. Астматикалық өкпедегі жасушадан тыс глутатион пероксидаза индукциясы: адамның тыныс алу жолдарының эпителий жасушаларында экспрессияның тотығу-тотықсыздануын реттеудің дәлелі. FASEB J. 2001;15:70�78.
74. Громер С, Уриг С, Беккер К. Тиоредоксин жүйесі ғылымнан клиникаға дейін. Med Res Rev. 2004;24:40�89.
75. Kinnula VL, Lehtonen S, Kaarteenaho-Wiik R, Lakari E, P��kk� P, et al. Адамның өкпесі мен өкпе саркоидозындағы пероксиредоксиндердің жасушаға тән экспрессиясы. Кеуде. 2002;57:157�164.
76. Дубуиссон М, Вандер Стрихт Д, Клиппе А, Этьен Ф, Наузер Т, т.б. Адамның пероксиредоксин 5 - пероксинитритредуктаза. FEBS Lett. 2004;571:161�165.
77. Холмгрен А. Тиоредоксин және глутаредоксин жүйелерінің антиоксиданттық қызметі. Антиоксидті тотығу-тотықсыздану сигналы. 2000;2:811�820.
78. Дикинсон Д.А., Форман ХДж. Қорғаныс және сигнал берудегі глутатион: кішкентай тиолдан алынған сабақтар. Энн NY Acad Sci. 2002;973:488�504.
79. Sies H. Глутатион және оның жасушалық қызметтегі рөлі. Free Radic Biol Med. 1999;27:916�921.
80. Ladner JE, Parsons JF, Rife CL, Gilliland GL, Armstrong RN. Глутатионтрансферазалардың параллельдік эволюциялық жолдары: rGSTK1-1 митохондриялық класс каппа ферментінің құрылымы мен механизмі. Биохимия. 2004;43:52�61.
81. Robinson A, Huttley GA, Booth HS, Board PG. Адамның каппа класы глутатионтрансферазасын модельдеу және биоинформатика зерттеулері прокариоттық 2-гидроксихром-2-карбоксилат изомеразаларына гомологиясы бар жаңа үшінші трансферазалар тобын болжайды. Biochem J. 2004;379:541�552.
82. Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hutchinson A, Persson B. MAPEGda жалпы құрылымдық ерекшеліктері эйкозаноид және глутатион метаболизмінде жоғары дивергентті функциялары бар мембранамен байланысты белоктардың кең таралған супертұқымы. Протеин ғылымы. 1999;8:689�692.
83. Хейс Дж.Д., Пулфорд диджейі. Глутатион S-трансфераза супергендік тобы: GST реттеуі және изоферменттердің қатерлі ісіктің химопротекциясына және дәріге төзімділігіне қосқан үлесі. Crit Rev Biochem Mol Biol. 1995;30:445-600.
84. Армстронг Р.Н. Глутатионтрансферазалардың құрылымы, каталитикалық механизмі және эволюциясы. Chem Res Toxicol. 1997;10:2�18.
85. Хейс Дж.Д., МакЛеллан Л.И. Глутатион және глутатионға тәуелді ферменттер тотығу стрессінен үйлестірілген реттелетін қорғанысты білдіреді. Free Radic Res. 1999;31:273�300.
86. Sheehan D, Meade G, Foley VM, Dowd CA. Глутатионтрансферазалардың құрылымы, қызметі және эволюциясы: ежелгі ферменттердің суперфамилиясының сүтқоректі емес мүшелерін жіктеуге салдары. Biochem J. 2001;360:1�16.
87. Чо С.Г., Ли ЮХ, Пак ХС, Рёо К, Канг КВ, т.б. Glutathione S-transferase Mu апоптоз сигналын реттейтін киназаны басу арқылы стресс белсендірілген сигналдарды модуляциялайды. J Biol Chem. 1;2001:276�12749.
88. Дорион С, Ламберт Н, Ландри Дж. Жылу соққысы арқылы p38 сигналдық жолын белсендіру Ask1-ден глутатион S-трансфераза Му диссоциациясын қамтиды. J Biol Chem. 2002;277:30792�30797.
89. Адлер V, Yin Z, Fuchs SY, Benezra M, Rosario L, т.б. GSTp арқылы JNK сигнализациясын реттеу. EMBO J. 1999;18:1321�1334.
90. Маневич Ю, Файнштейн С.И., Фишер А.Б. Антиоксидантты фермент 1-CYS пероксиредоксиннің белсендірілуі pGST-пен гетеродимеризация арқылы жүргізілетін глутатионилденуді қажет етеді. Proc Natl Acad Sci US A. 2004;101:3780�3785.
91. Бункер VW. Бос радикалдар, антиоксиданттар және қартаю. Med Lab Sci. 1992;49:299�312.
92. Mezzetti A, Lapenna D, Romano F, Costantini F, Pierdomenico SD және т.б. Жүйелік тотығу стрессі және оның жас және аурумен байланысы. J Am Geriatr Soc. 1996;44:823�827.
93. Уайт Е, Шеннон Дж.С., Паттерсон Р.Е. Витамин мен арасындағы байланыс
кальций қоспаларын қолдану және тоқ ішек ісігі. Қатерлі ісік эпидемиолын биомаркерлері Алдыңғы. 1997;6:769-774.
94. Masella R, Di Benedetto R, Vari R, Filesi C, Giovannini C. Биологиялық жүйелердегі табиғи антиоксиданттық қосылыстардың жаңа механизмдері: глутатион мен глутатионмен байланысты ферменттердің қатысуы. J Nutr Biochem. 2005;16:577�586.
95. Curello S, Ceconi C, Bigoli C, Ferrari R, Albertini A, Guarnieri C. Ишемия мен реперфузиядан кейінгі жүрек глутатионының күйіндегі өзгерістер. Тәжірибе. 1985;41:42�43.
96. Эль-Агамей А, Лоу ГМ, МакГарви диджей, Мортенсен А, Филлип ДМ, Трускотт Т.Г. Каротиноидтардың радикалды химиясы және антиоксиданттық/прооксиданттық қасиеттері. Арқа биохимиясының биофизи. 2004;430:37�48.
97. Райс-Эванс CA, Sampson J, Bramley PM, Holloway DE. Неліктен біз каротиноидтарды in vivo антиоксиданттар деп күтеміз? Free Radic Res. 1997;26:381�398.
98. Найлс Р.М. Ретиноидты химиопрофилактика және қатерлі ісіктерді емдеудегі сигналдық жолдар. Мутат Рес. 2004;555:81�96.
99. Донато Л.Д., Ной Н. Сүт безінің карциномасының өсуін ретиной қышқылы арқылы басу: проапоптотикалық гендер ретиной қышқылы рецепторлары мен жасушалық ретиноин қышқылын байланыстыратын ақуыз II сигналының нысанасы болып табылады. Cancer Res. 2005;65:8193�8199.
100. Ниизума Х, Накамура Ю, Озаки Т, Наканиши Х, Охира М және т.б. Bcl-2 нейробластомадағы ретиноин қышқылының әсерінен апоптозды жасуша өлімінің негізгі реттеушісі болып табылады. Онкоген. 2006;25:5046�5055.
101. Дальтон Т.П., Шерцер Х.Г., Пуга А. Реактивті оттегі арқылы ген экспрессиясының реттелуі. Энн Рев фармакол токсиколы. 1999;39:67�101.
102. Scandalios JG. Тотығу стрессіне геномдық жауаптар. In: Meyers RA, ed. Молекулярлық жасуша биологиясы және молекулалық медицина энциклопедиясы. 5-том. 2-ші басылым. Weinheim, Германия: Wiley-VCH; 2004: 489-512.
103. Гош Р, Митчелл Д.Л. Промоторлық элементтердегі тотығу ДНҚ зақымдануының транскрипция факторымен байланысуына әсері. Нуклеин қышқылдары Res. 1999;27:3213�3218.
104. Мариетта Си, Гулам Х, Брукс ПДЖ. TATA қорабындағы жалғыз 8, 50-цикло-20-дезоксиаденозин зақымдануы TATA байланыстыратын ақуызды байланыстыруды болдырмайды және in vivo транскрипциясын қатты төмендетеді. ДНҚ жөндеу (Amst). 2002;1:967-975.
105. Джексон АЛ, Чен Р, Лоеб ЛА. Микросателлиттік тұрақсыздықтың индукциясы
тотықтырғыш ДНҚ зақымдануымен. Proc Natl Acad Sci US A. 1998;95:12468-12473.
106. Кальдекот КВт. Сүтқоректілердің ДНҚ-ның бір тізбекті үзілуін қалпына келтіру кезінде белок пен ақуыздың өзара әрекеттесуі. Biochem Soc Trans. 2003;31:247�251.
107. Cooke MS, Evans MD, Dizdaroglu M, Lunec J. ДНҚ-ның тотығу зақымдалуы: механизмдер, мутация және ауру. FASEB J. 2003;17:1195-1214.
108. Джонс П.Л., Вольфф AP. Геннің экспрессиясын анықтаудағы хроматинді ұйымдастыру мен ДНҚ метилденуі арасындағы байланыстар. Semin Cancer Biol. 1999;9:339�347.
109. Жиротти AW. Липидтердің асқын тотығу механизмдері. J Free Radic Biol Med. 1985;1:87-95.
110. Сиу Г.М., Дрейпер Х.Х. Малональдегидтің in vivo және in vitro метаболизмі. Липидтер. 1982;17:349-355.
111. Эстербауэр Х, Коллер Е, Сли Р.Г., Костер Дж.Ф. Флуоресцентті хромолипидтердің түзілуіне 4-гидроксиноненалдың липидті-тотықсыздану өнімінің мүмкін қатысуы. Biochem J. 1986;239:405�409.
112. Хагихара М, Нишигаки I, Масеки М, Яги К. Адам сарысуының липопротеиндік фракцияларындағы липидті асқын тотығы деңгейіндегі жасқа байланысты өзгерістер. Дж Геронтол. 1984;39:269�272.
113. Келлер Дж.Н., Марк РДЖ, Брюс А.Дж., Бланк Е, Ротштейн Дж.Д., т.б. 4- Гидроксиноненал, мембраналық липидтердің асқын тотығуының альдегидтік өнімі, синаптосомалардағы глутаматтың тасымалдануын және митохондриялық қызметін бұзады. Неврология. 1997;806:85�96.
114. Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. Липидтердің асқын тотығуының соңғы өнімі арқылы стресс сигнал беру жолдарын белсендіру. 4-гидрокси-2-ноненал жасушаішілік пероксид өндірісінің потенциалды индукторы болып табылады. J Biol Chem. 1999;274:2234�2242.
115. Suc I, Meilhac O, Lajoie-Mazenc I, Vandaele J, Jurgens G, Salvayre R, Negre-Salvayre A. EGF рецепторын тотыққан LDL арқылы белсендіру. FASEB J. 1998;12:665-671.
116. Цукагоши Н, Кавата Т, Шимизу Ю, Ишизука Т, Добаши К, Мори М. 4-Гидрокси-2-ноненал адамның өкпе фибробласттары IMR-90 арқылы фибронектин өндірісін ішінара эпидермальды өсу факторы рецепторымен байланысты жасушадан тыс сигналды белсендіру арқылы күшейтеді. реттелетін киназа p44/42 жолы. Toxicol Appl Pharmacol. 2002;184:127�135.
117. Montuschi P, Collins JV, Ciabattoni G, Lazzeri N, Corradi M, Haritonov SA, Barnes PJ. ӨСОА бар емделушілерде және дені сау темекі шегушілерде өкпенің тотығу стрессінің in vivo биомаркері ретінде дем шығаратын 8-изопростан. Am J Respir Crit Care Med. 2000;162:1175�1177.
118. Моррисон Д, Рахман I, Ланнан С, МакНи В. Шылым шегушілердің ауа кеңістігіндегі эпителий өткізгіштігі, қабыну және тотықтырғыш стресс. Am J Respir Crit Care Med. 1999;159:473�479.
119. Новак Д, Касиельски М, Антчак А, Пиетрас Т, Биаласевич П. Тұрақты созылмалы обструктивті өкпе ауруы бар науқастардың тыныс алу конденсатында тиобарбитур қышқылы-реактивті заттардың және сутегі асқын тотығының жоғарылауы: темекі шегудің айтарлықтай әсері жоқ. Respir Med. 1999;93:389�396.
120. Келли ФДЖ, Мудвэй IS. Ауа-өкпе интерфейсінде ақуыздың тотығуы. Амин қышқылдары. 2003;25:375�396.
121. Дин Р.Т., Робертс CR, Джессуп В. Жасушадан тыс және жасушаішілік полипептидтердің бос радикалдармен фрагментациясы. Prog Clin Biol Res. 1985;180:341�350.
122. Кек Р.Г. Белоктардағы метиониннің тотығуы үшін зонд ретінде т-бутил гидропероксидін қолдану. Анальды биохимия. 1996;236:56�62.
123. Дэвис КДж. Ақуыздың зақымдануы және оттегі радикалдарымен ыдырауы. I. Жалпы аспектілер. J Biol Chem. 1987;262:9895�9901.
124. Stadtman ER. Белоктардың металл иондарымен катализделген тотығуы: биохимиялық механизмі және биологиялық салдары. Free Radic Biol Med.
1990; 9: 315 325.
125. Fucci L, Oliver CN, Coon MJ, Stadtman ER. Аралас функциялы тотығу реакциялары арқылы негізгі метаболикалық ферменттерді инактивациялау: ақуыз айналымы мен қартаюға ықтимал салдары. Proc Natl Acad Sci US A. 1983;80:1521-1525.
126. Stadtman ER, Moskovitz J, Levine RL. Белоктардың метионин қалдықтарының тотығуы: биологиялық салдары. Антиоксидті тотығу-тотықсыздану сигналы. 2003;5:577�582.
127. Stadtman ER, Levine RL. Белоктардағы бос аминқышқылдарының және аминқышқылдарының қалдықтарының бос радикалдар арқылы тотығуы. Амин қышқылдары. 2003;25:207�218.
128. Stadtman ER. Қартаю және қартаюға байланысты ауруларда ақуыздың тотығуы. Энн NY Acad Sci. 2001;928:22�38.
129. Shacter E. Биологиялық үлгілердегі ақуыздың тотығуының сандық көрсеткіші және маңызы. Drug Metab Rev. 2000;32:307�326.
130. Поли Г, Леонардузци Г, Биаси Ф, Чиарпотто Е. Тотығу стрессі және жасуша сигнализациясы. Curr Med Chem. 2004;11:1163-1182.
131. Нойфельд Г, Коэн Т, Генгринович С, Полторак З. Тамырлық эндотелий өсу факторы (VEGF) және оның рецепторлары. FASEB J. 1999;13:9�22.
132. Sundaresan M, Yu ZX, Ferrans VJ, Sulciner DJ, Gutkind JS, т.б. Rac1 бойынша фибробласттарда реактивті-оттегі түрлерінің генерациясын реттеу. Biochem J. 1996;318:379�382.
133. Sun T, Oberley LW. Транскрипциялық активаторлардың тотығу-тотықсыздану реттелуі. Free Radic Biol Med. 1996;21:335�348.
134. Klatt P, Molina EP, De Lacoba MG, Padilla CA, Martinez-Galesteo E, Barcena JA, Lamas S. Қайтымды S-глутатиоляция арқылы c-Jun ДНҚ байланысуының тотықсыздандырылуын реттеу. FASEB J. 1999;13:1481-1490.
135. Reynaert NL, Ckless K, Guala AS, Wouters EF, van der Vliet A, Janssen Heininger
YM. Глутаредоксин-1 катализделген цистеин деривациясынан кейін S-глутатионилденген ақуыздарды in situ анықтау. Biochim Biophys Acta. 2006;1760:380�387.
136. Reynaert NL, Wouters EF, Janssen-Heininger YM. Глутаредоксин-1 модуляциясы
тыныс жолдарының аллергиялық ауруларының тышқан үлгісіндегі экспрессиясы. Am J Respir Cell Mol Biol. 2007;36:147�151.
137. Филомени Г, Ротилио Г, Цириоло MR. Жасуша сигнализациясы және глутатион тотығу-тотықсыздану жүйесі. Biochem Pharmacol. 2002;64:1057�1064.
138. Панде В, Рамос М.Дж. 15-дезоксидельта (12,14) простагландин J(2) ядролық фактор-каппа В және басқа жасушалық ақуыздар арқылы молекулалық тану. Bioorg Med Chem Lett. 2005;15:4057�4063.
139. Перкинс Н.Д. NF-kappaB және IKK функциясымен ұяшықты сигнал беру жолдарын біріктіру. Nat Rev Mol Cell Biol. 2007;8:49�62.
140. Гилмор Т.Д. NF-kappaB-ке кіріспе: ойыншылар, жолдар, перспективалар. Онкоген. 2006;25:6680�6684.
141. Хирота К, Мурата М, Сачи Ю, Накамура Х, Такеучи Дж, Мори К, Йодой Дж. Тиоредоксиннің цитоплазмадағы және ядродағы ерекше рөлдері. NF-kappaB транскрипция факторының тотығу-тотықсыздану реттелуінің екі сатылы механизмі. J Biol Chem. 1999;274:27891�27897.
142. Палата ПА. Өкпенің жедел зақымдалуындағы комплементтің, хемокиндердің және реттеуші цитокиндердің рөлі. Энн NY Acad Sci. 1996;796:104�112.
143. Akira S, Kishimoto A. NF-IL6 және NF-kB цитокиндік генді реттеу. Adv Иммунол. 1997;65:1�46.
144. Мейер М, Шрек Р, Бауэрле П.А. H2O2 және антиоксиданттар NF-kappa B және AP-1 интактілі жасушаларда белсендіруге қарама-қарсы әсер етеді: AP-1 екінші антиоксидантқа жауап беретін фактор ретінде. EMBO J. 1993;12:2005�2015.
145. Abate C, Patel L, Rausher FJ, Curran T. in vitro фос және jun ДНҚ-байланыстыру белсенділігінің тотықсыздандырылуын реттеу. Ғылым. 1990;249:1157�1161.
146. Galter D, Mihm S, Droge W. Дисульфид глутатионының кВ ядролық транскрипция факторларына және белсендіруші ақуыз-1 әсерлері. Eur J Biochem. 1994;221:639�648.
147. Хирота К, Мацуи М, Ивата С, Нишияма А, Мори К, Йодой Дж. AP-1 транскрипциялық белсенділігі тиоредоксин мен Реф-1 арасындағы тікелей байланыс арқылы реттеледі. Proc Natl Acad Sci US A. 1997;94: 3633�3638.
IFM's Find A Practitioner құралы пациенттерге әлемнің кез келген жерінде функционалдық медицина мамандарын табуға көмектесу үшін жасалған функционалдық медицинадағы ең үлкен бағыттаушы желі болып табылады. IFM сертификатталған тәжірибешілері олардың функционалдық медицина бойынша кең білімін ескере отырып, іздеу нәтижелерінде бірінші болып табылады.